2015-04-06
2015-04-06 887
887В аутентичной неживой природе (исключающей технику) на макроскопическом уровне информация выражается, как мы выяснили, в мере свободной энергии – то есть в способности системы совершить некоторую работу – механическую, химическую, электромагнитную и т.д. Каждый процесс такого совершения работы сопровождается необратимой (без внешнего воздействия) потерей свободной энергии, а значит, как мы вывели до этого, и информации. В случае внешнего воздействия некоторая система может получить долю свободной энергии, израсходованной в системе, внешней по отношению к ней, при этом возникает такое состояние частиц, которое позволяет системе совершить работу. Классический пример – намагничивание ферромагнетиков либо другим магнитом, либо ударом молнии. Намагниченность в данном случае будет физическим выражением информации, полученной частицами данной системы. Работу в данном случае будет совершать магнитное поле. Процессы передачи, хранения и использования информации в данном случае – одноступенчатые и не имеющие цели. Еще одна особенность информационных воздействий в неживой природе – при передаче информации от одной системы к другой возрастание информации в системе-приемнике необходимо сопряжено с ее потерей в системе-источнике.
В связи с развитием квантовой электроники представляет интерес рассмотрение информационных процессов в мире элементарных частиц. Каждая частица имеет либо массу покоя (эквивалентную общей энергии), либо энергию без массы покоя. Кроме того, у каждой частицы есть стандартный набор дискретных характеристик – квантовых чисел, одни из которых собственно характеризуют ее неизменные свойства, присущие ей все время ее существования, другие зависят от полученных-отданных квантов энергии и способны характеризовать ее текущее состояние (энергетический уровень). Таким образом, каждая элементарная частица имеет ряд дискретных степеней свободы, а значит – является носителем некоторого количества информации. При взаимодействии частиц, сопровождающемся их превращениями, имеется ряд законов сохранения – энергии, барионного квантового числа, лептонного квантового числа, и некоторые другие. При этом фундаментальные квантовые числа меняются определенным образом для каждого взаимодействия, а это значит, что к взаимодействию частиц применимо понятие марковских подстановочных алгоритмов. Энтропии в классическом понимании в мире элементарных частиц не существует, но необратимые потери свободной энергии происходят – например, потеря энергии с антинейтрино при распаде свободного нейтрона на протон и электрон. Таким образом, закон невозрастания информации для мира элементарных частиц справедлив. Для мира элементарных частиц в отличие от макроскопического существует еще она принципиальная особенность – невозможность хранения информации в случае количества частиц порядка единиц. Каждая частица при изменении своего состояния не оставляет никакого следа о своих прошлых состояниях.
Учитывая следующие факторы:
1. Кроме квантовых чисел и энергетических уровней (т.е. степеней свободы) элементарных частиц нам о них неизвестно больше ничего;
2. Элементарные частицы при взаимодействии переходят друг в друга по определенным алгоритмам;
3. Согласно релятивистскому подходу существует эквивалентность массы и энергии, выраженная в формуле Эйнштейна E=mc2;
- можно заключить, что вся материя является формой существования энергии, которая дискретизирована в структуры элементарных частиц, то есть структурирована при помощи информации. Таким образом, все разнообразие материального мира – это форма существования одной субстанции, при помощи информации (т.е. набора дискретных параметров) принимающей все возможные формы.
В живой природе мы видим знакомые нам по вычислительной технике схемы работы с информацией. Имеется источник информации – генетический материал в виде ДНК и РНК. Имеется присущий этому источнику четырехбуквенный генетический код, состоящий из букв -мононуклеотидов А, Г, Т, Ц (аденин, гуанин, тимин, цитозин), который в РНК модифицирован – тимин заменен на урацил. Буквы эти сочетаются в трехбуквенные слова (триплеты), которые являются кодом для синтеза аминокислот (3 буквы соответствуют или аминокислоте, или же «пустому» слову, означающему прерывание синтеза). Имеется кодирующее устройство – РНК-полимераза, которое создает промежуточное сообщение в виде информационной РНК для декодирующего устройства – рибосомы, которое на основании полученного сообщения синтезирует полипептидные цепочки, которые потом, изменяя конформацию уже без участия генетического источника информации, складываются в молекулы белков. Все эти процессы идут с затратой свободной энергии, запасенной в виде макроэргических связей веществ, их образующих (АТФ, ГТФ, креатинфосфат и некоторые другие). Сама же свободная энергия запасается или при захвате фотонов при фотосинтезе у автотрофов (растений), или же при окислении органических субстратов у гетеротрофов (животных, грибов, бактерий и т.д.). И тот, и другой процессы осуществляются при помощи ферментативных реакций (реакций с участием ферментов, т.е. простых или составных белков, являющихся катализаторами – хотя роль ферментов гораздо сложнее обычного химического катализа, как мы это рассмотрим позже) как правило, многоступенчатых, с последовательным участием различных ферментов, синтезированных на основе имеющегося генетического кода. Кроме белков, участвующих в энергетическом обмене, существует множество регуляторных ферментов, осуществляющих функции обратных связей, которые регулируют как экспрессию генов и синтез белков и других органических соединений, так и энергетический обмен. Кроме того, генетическая информация является самокопирующейся – как внутри клетки при помощи комплементарного преобразования (перекодирования) А-Т и Г-Ц, осуществляющегося при синтезе комплементарной спирали ДНК, которая и хранится в виде двойной спирали, так и при процессах деления клетки (митоз и мейоз), при этом осуществляется репликация ДНК в новые двойные спирали, предназначенные для хранения и считывания в дочерних клетках.
Итак, мы видим в живой природе по сравнению с неживой множество новых особенностей существования информации.
1. Важным является не только количество, но и качественный состав информации – последовательность ДНК кодирует через синтез ферментов процессы, которые эти ферменты осуществляют. То есть каждый фермент является выраженным в виде химического соединения оператором, осуществляющим химические же преобразования веществ в цитоплазме клетки.
2. Ферменты, участвующие в биохимических преобразованиях, функционально соединены в последовательности, реализующие некую конечную цель (синтез определенного продукта), то есть каждый биохимический цикл является выражением некоего алгоритма. Эти алгоритмы надо понимать не в смысле Тьюринга, а в более общем смысле Маркова – то есть это подстановочные алгоритмы, осуществляющие преобразования в «словах» - органических соединениях (или меняющие концентрацию неорганических, как в водно-электролитном обмене).
3. При наличии в этих циклах регуляторных ферментов (или аллостерических регуляторных центров, стимулирующих или ингибирующих активный центр фермента при соединении с субстратом или продуктом реакции – или же сторонним регулирующим химическим агентом) эти алгоритмы являются циклическими, осуществляющими регуляцию по типу обратной связи.
4. В связи с диффузным расположением молекул субстратов в растворе биохимические циклы-алгоритмы являются не абсолютно детерминированными, а вероятностными, причем вероятность осуществления следующей ступени алгоритма зависит от концентрации субстрата. То есть данные алгоритмы по своей реализации представляют из себя марковские цепи.
Вероятностность биохимических алгоритмов позволяет описать их в категориях шенноновской теории информации. Итак, биохимический алгоритм есть процесс, состоящий из последовательности информационных воздействий при посредстве биохимических агентов и увеличивающий вероятность возникновения конкретного состояния-цели в биохимической системе при ступенчатом повышении условной вероятности данного состояния по сравнению с безусловной. Под биохимической системой можно понимать как отдельные клеточные органеллы, например, митохондрии, так и целые клетки и организмы, и даже сложные надорганизменные системы типа системы «самец-самка» в брачный период, поведение которой управляется выделяемыми во внешнюю среду информационными биохимическими агентами – феромонами. Каждая ступень биохимического алгоритма, повышающая вероятность состояния-цели, является конкретной биохимической реакцией, использующей продукт предыдущей ступени в качестве субстрата для настоящей, а продукт настоящей реакции будет являться субстратом для последующей ступени – и так до конечной ступени, синтезирующей конечный продукт.
Интересно совпадение трактовки «машина-алгоритм» в абстрактных моделях Тьюринга, Поста, Черча (для них эти термины были синонимами), с сущностью биохимических процессов. Ферментная система является биохимической машиной, работающей за счет конформационных изменений (фактически, из-за участия в этих изменениях состояний электронных оболочек – квантово-механической конструкцией), и одновременно – она является совокупностью сообщений для молекул субстрата о том, какие изменения должны быть над ними произведены. Биохимическая машина не просто выполняет алгоритмы, а сама является материализованным алгоритмом, записанным на языке химических соединений.
Конечной целью всех информационных процессов в живом организме является передача генетической информации следующему поколению. Для реализации этой цели в биохимических (а также почти не рассматриваемых здесь поведенческих) алгоритмах предусмотрены промежуточные цели:
1. Обеспечение оптимальных условий хранения генетической информации, выражающееся в стремлении к постоянству внутренней среды клетки, которое называется гомеостаз. Цель гомеостаза – максимально противостоять разрушению генетического материала, то есть минимизировать действие закона невозрастания информации.
2. Обеспечение копирования генетической информации как внутри организма (геном каждой клетки в идеале является точной копией соседних), так и при передаче следующему поколению при бесполом или половом размножении.
3. Обеспечение рекомбинации генетической информации при половом размножении при помощи механизма кроссинговера (обмена генами между гомологичными хромосомами) для получения наибольшего генетического разнообразия у следующего поколения. Кроссинговер по своей сути – тоже марковский подстановочный алгоритм.
4. Обеспечение реакций организма на изменения внешней среды, которые потенциально могут быть или полезными для вышеуказанных целей, или же опасными. Для примитивных организмов это чисто биохимические или биофизические алгоритмы – тропизмы и настии, для животных, имеющих нервную систему – поведенческие алгоритмы. Изначально заложенные в нервной системе поведенческие алгоритмы называются безусловными рефлексами, появившиеся в результате многократного реагирования на внешние стимулы – условными.
Ценность (то есть численное выражение важности его смысла) какого-либо сообщения или алгоритма по Корогодину (см.) можно записать в виде простой формулы:
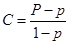 ,
,
где P – условная вероятность появления состояния-цели при реализации алгоритма, а p – ее безусловная вероятность.
Идеальный алгоритм делает условную вероятность P®1, и тогда C®1, бесполезный алгоритм – при нем P=p, и тогда C=0, но возможен и вариант, когда P<p, при этом C<0, и тогда данное информационное воздействие можно назвать дезинформацией.
Но существует и другой критерий оценки ценности информации, позволяющий непосредственно оценивать ее в информационных (или энтропийных) единицах. Это формула Харкевича (см.):
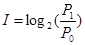 ,
,
где P1 – условная вероятность состояния-цели при информационном воздействии, P0 – безусловная вероятность достижения этого состояния. В отличие от нормированной оценки по Корогодину, этот вариант позволяет произвести количественную оценку ценности информации, напрямую измеряя ее в битах.
Дезинформирующими агентами можно считать любые воздействия, нарушающие процесс хранения и передачи генетической информации и реализации биохимических алгоритмов. На генном уровне это радиация и химические мутагены, на субклеточном уровне – яды, свободные радикалы, нарушения водно-электролитного обмена и многие другие. Особого внимания заслуживают вирусы – это также системы хранения и передачи генетической информации, которые тратят пластические и энергетические ресурсы и механизмы экспрессии генов хозяина, тем самым создавая помехи для его информационных процессов. Некоторые вирусы способны встраивать свою ДНК в геном хозяина, тем самым увеличивая количество информации в его ДНК – то есть здесь мы имеем дело с настоящим информационным воздействием извне. При этом семантическая ценность информации ДНК хозяина в общем случае снижается, но в случае, если вирус уже побывал у другого хозяина и захватил в свой геном его гены, а затем эти гены передает другому хозяину, возможно добавление новой информации, имеющей семантическую ценность. Это явление называется горизонтальным переносом генов.
Закон невозрастания информации работает не только на генетическом уровне, но и на уровне систем обеспечения гомеостаза, что ведет к накоплению ошибок в биохимических алгоритмах и снижению семантической ценности последних, а значит – к все меньшей вероятности достижения состояния-цели (гомеостаза). При некотором критическом накоплении ошибок достижение гомеостаза становится невозможным (алгоритмы становятся бесполезными), и необратимые изменения внутренней среды приводят к смерти организма.
Представляет большой интерес количественная оценка информации в живых системах. Согласно выводам Л.А. Блюменфельда каждый человеческий организм содержит около 2,6*1022 бит информации, что в принципе не превышает, по его же оценке, величину информации в куске горной породы той же массы – то есть говорить о принципиальном количественном отличии живого от неживого по этой оценке бессмысленно. Но по его же заявлению, это вызывает вопрос о некорректности такого критерия. Действительно, в оценке этих различий нет временного критерия информации, и поэтому оценить отличие поможет введение потока информации, то есть количества обрабатывающейся информации на какую-либо биологическую единицу (ферментную систему, клетку, организм) за единицу времени. Этим мы сейчас и займемся.
Итак, при работе ферментной системы происходит увеличение вероятности продуктов реакции от исчезающе малой (по оценкам того же Блюменфельда около 10-23) до очень большой – где-то 0,6 – 0,7. Количество информации за один такт работы ферментной системы будет равно:
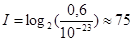 бит.
бит.
Длительность одного такта работы фермента – около 10-6 с, то есть поток информации при работе одного фермента равен 75*106 бит/с. Количество белков в одной клетке порядка 108, и поскольку большинство из них является ферментами, поток информации через одну клетку будет около 75*1014 бит/с. Количество клеток в организме человека около 1013, таким образом, поток информации на одного живого человека составит 75*1027 бит/с. Эти расчеты, разумеется, не точны и позволяют оценить только порядок скорости обработки информации в живом организме. Разумеется, в данном случае кардинальное отличие живого организма от куска неживой материи (в том числе и от свежего трупа) очевидно, и «парадокс Блюменфельда» можно считать разрешенным.
Выводы:
1. Информационные процессы в неживой природе одноступенчаты, необратимы, сопровождаются потерей информации в системе-источнике при ее получении системой-приемником. Закон невозрастания информации справедлив как для макроскопических неживых систем, так и для мира элементарных частиц.
2. Кардинальным различием информационных процессов в неживых системах (без участия техники) и живых организмах является отсутствие состояний-целей в первом случае и их наличие во втором. Для каждого живого организма существует конечная цель – передача генетической информации следующим поколениям, и промежуточные цели – обеспечение хранения и передачи этой информации при помощи биохимических, а в случае животных – и поведенческих алгоритмов. Хранение генетической информации в макромолекулах показывает уровень, начиная с которого возможно хранение информации вообще – это граница между молекулярным и макроскопическим уровнями, т.е. минимальная единица хранения информации – это макромолекула.
3. Для реализации успешного хранения генетической информации в живых организмах имеются механизмы гомеостаза – обеспечения постоянного стремления к некоторому благоприятному для этого состоянию-цели, при котором действие закона невозрастания информации минимально, а соответственно – минимальны потери генетической информации. Эти механизмы обеспечиваются множеством биохимических и поведенческих (у животных) алгоритмов, которые снабжены механизмами обратной связи.
4. Как биохимические, так и поведенческие алгоритмы способны изменяться при устойчивых изменениях внешней среды, которые дают информацию для систем обратной связи. Этот процесс можно назвать обучаемостью живых систем.
5. Генетическая и биохимическая информация имеет не только количественную характеристику, но и качественную – семантическую ценность, которая выражается в увеличении условной вероятности достижения состояния-цели (гомеостаза). Эта ценность может быть максимальной в идеале, а может снижаться при наличии дезинформирующх агентов.
6. Смерть живого организма наступает при некотором критическом снижении семантической ценности алгоритмов обеспечения гомеостаза вследствие действия закона невозрастания информации (т.е. накопления алгоритмических ошибок), при котором они становятся бесполезными и приводят к изменениям внутренней среды, несовместимым с достижением состояния-цели.
7. Кардинальное отличие в количественной оценке живых и неживых систем определяется не собственно количеством информации, а ее потоком. Поток информации в неживых системах фактически нулевой, в живых же характеризуется величинами порядка от 1014 бит/с (единичные клетки и одноклеточные организмы) до 1027 бит/с (организм в целом). Смерть организма является прекращением потока информации.