 2015-04-30
2015-04-30 1111
1111Необратимое ингибирование наблюдают в случае образования ковалентных стабильных связей между молекулой ингибитора и фермента. Чаще всего модификации подвергается активный центр фермента, В результате фермент не может выполнять каталитическую функцию.
К необратимым ингибиторам относят ионы тяжёлых металлов, например ртути (Hg2+), серебра (Ag+) и мышьяка (As3+), которые в малых концентрациях блокируют сульфгидрильные группы активного центра. Субстрат при этом не может подвергаться химическому превращению (рис. 2-26). При наличии реактиваторов ферментативная функция восстанавливается. В больших концентрациях ионы тяжёлых металлов вызывают денатурацию белковой молекулы фермента, т.е. приводят к полной инактивации фермента.
1. Специфические и неспецифические
ингибиторы
Использование необратимых ингибиторов представляет большой интерес для выяснения
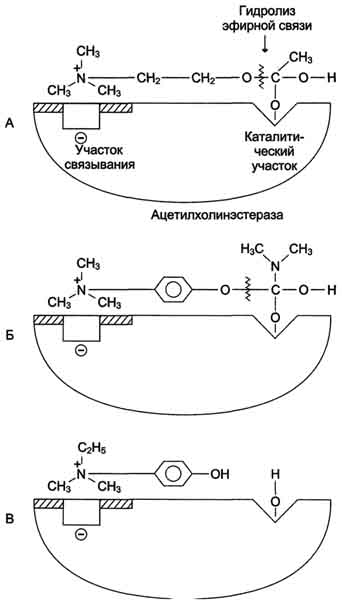
Рис. 2-23. Схема активного центра ацетилхолинэстеразы. А - присоединение ацетилхолина в активном центре фермента. Стрелкой указано место гидролиза эфирной связи в молекуле ацетилхолина; Б - присоединение конкурентного ингибитора - прозерина в активном центре фермента. Указано место гидролиза прозерина, однако реакция идёт намного медленнее, чем с ацетилхолином; В - присоединение конкурентного ингибитора в активном центре фермента - эндрофония. Эндрофоний связывается в активном центре ацетилхолинэстеразы, препятствуя присоединению ацетилхолина.
механизма действия ферментов. С этой целью применяют вещества, блокирующие определённые группы активного центра ферментов. Такие ингибиторы называют специфическими. Ряд соединений легко вступает в реакции с определенными
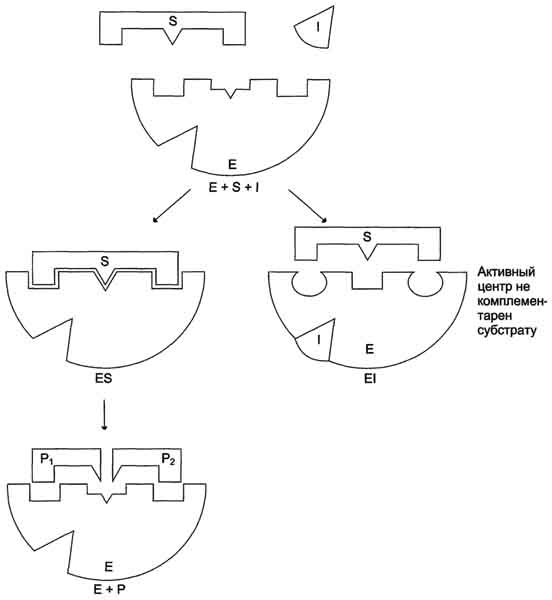
Рис. 2-24. Схема неконкурентного ингибирования активности фермента.
химическими группами. Если эти группы участвуют в катализе, то происходит полная инактивация фермента.
Роль гидроксильных групп серина в механизме катализа исследуют с помощью фторфосфатов, например диизопропилфторфосфата. Дии-зопропилфторфосфат (ДФФ) специфически реагирует лишь с одним из многих остатков серина в активном центре фермента. Остаток Сер, способный реагировать с ДФФ, имеет идентичное или очень сходное аминокислотное окружение (табл. 2-2). Высокая реакционная способность этого остатка по сравнению с другими остатками Сер обусловлена аминокислотными остатками, также входящими в активный центр ферментов.
ДФФ относят к специфическим необратимым ингибиторам "сериновых" ферментов, так как он образует ковалентную связь с гидроксильной группой серина, находящегося в активном центре и играющего ключевую роль в процессе катализа (рис. 2-27).
Ацетат йода, п-хлормеркурибензоат легко вступают в реакции с SH-группами остатков цистеина белков (рис. 2-28). Эти ингибиторы не относят к специфичным, так как они
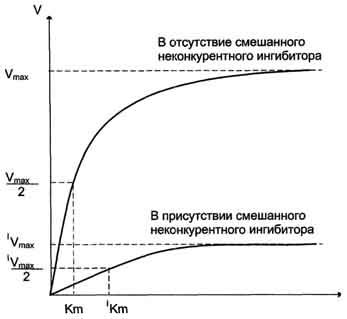
Рис. 2-25. Влияние неконкурентного ингибитора на скорость ферментативной реакции в зависимости от концентрации субстрата. Vmax - максимальная скорость реакции в отсутствие ингибитора; 'Vmax - максимальная скорость реакции в присутствии ингибитора; Кm - константа Михаэлиса в отсутствие ингибитора; 'Кm - константа Михаэлиса в присутствии ингибитора.
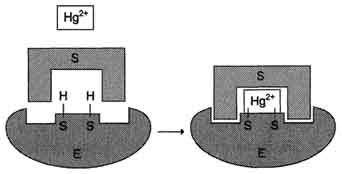
Рис. 2-26. Механизм действия ионов ртути как необратимого ингибитора. Ионы ртути в малых концентрациях блокируют сульфгидрильные группы активного центра, что приводит к снижению скорости ферментативной реакции.
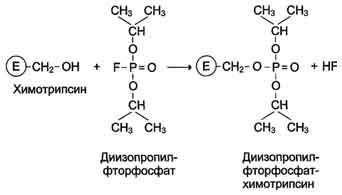
Рис. 2-27. Ингибирование активности химотрипсина с помощью диизопропилфторфосфата.

