 2015-06-26
2015-06-26 528
528Вопрос о последовательности событий, приводящих к запуску ответной реакции растения на О2-дефицит, и о природе рецептора сигнала до настоящего времени остается нерешенным. В анаэробных условиях быстро снижается уровень АТФ, увеличивается содержание НАДН (НАДФН) и происходит закисление цитозоля. Любой из этих факторов может принимать участие в процессах сигнальной трансдукции. Как будет показано ниже, в пути передачи сигналов, отвечающих за образование аэренхимы при О2-дефиците, вовлечен гормон этилен. Возможно, что этилен, а также АБК участвуют в запуске и других процессов, индуцируемых О2-дефицитом.
Вторичным мессенджером в путях сигнальной трансдукции, активируемых аноксией, является
Са2+: его содержание в протопластах кукурузы при анаэробных условиях возрастает. Повышенные концентрации Са2+ необходимы для инициации транскрипции гена алкогольдегидрогеназы Adh 1.
При аноксии в корнях проростков кукурузы происходят существенные изменения в экспрессии генов и метаболизме. Они обнаруживаются уже через несколько часов после создания анаэробных условий. Общий синтез белков подавляется на 70 %. Гипоксия приводит к аналогичному эффекту, однако он выражен в значительно меньшей степени.
Изменения в синтезе белка при аноксии связаны с регуляцией экспрессии генов на транскрипционном и посттранскрипционном уровнях. Большая часть генов, активных в присутствии О2, при аноксии репрессируется. В то же время некоторый набор генов, не экспрессирующихся до создания анаэробных условий, в условиях аноксии активируется. Экспрессия этих генов чрезвычайно важна для выживания растений в бескислородных условиях. Подавляющая их часть относится к генам ферментов, которые осуществляют (см. подразд. 8.5.3) процессы деградации крахмала, гликолиза и ферментации этанола. Некоторые из этих ферментов, в частности АДГ и глицеральдегид-3-фосфатдегидрогеназа, представлены в клетках несколькими изоформами. Одни изоформы образуются в клетках в аэробных условиях, т. е. характеризуются конститутивным синтезом и участвуют в нормальном метаболизме, другие индуцируются в ответ на О2-дефицит. Гены, кодирующие эти две группы изоформ, различаются по нуклеотидным последовательностям промоторов. Гены, экспрессирующиеся в ответ на О2-дефицит, называются анаэробными генами, а их продукты анаэробными белками. Некоторые анаэробные гены клонированы, в частности гены алкогольдегидрогеназы Adh из кукурузы и арабидопсиса, а также гены пируватдекарбоксилазы Pdc из арабидопсиса. Были охарактеризованы гены Adh. Их про-моторные области содержат специфические цис- элементы, которые необходимы для экспрессии в условиях кислородного дефицита (рис. 8.31).
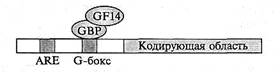
Рис. 8.31. Гены алкогольдегидрогеназы АДГ из кукурузы и Arabidopsis.
Эти гены содержат в промоторной области специфические цис -элементы, которые необходимы для экспрессии в анаэробных условиях, — ARE (anaerobic response element) и G-бокс. Два транскрипционных фактора взаимодействуют с G-боксом гена Adh 1 из Arabidopsis: GBP (G-box-binding factor) и GF14 (G-factor). Для индукции транскрипции GF14 должен сформировать комплекс с GBP, чтобы последний мог вступить в непосредственное взаимодействие с нуклеотидной последовательностью G-бокса
Известны две такие специфические последовательности: ARE (anaerobic response element) и так называемый G-бокс. Идентифицированы два транскрипционных фактора, взаимодействующих с G-боксом гена Adh 1 из арабидопсиса: GBP (G-box-binding protein) и GF14 (G-factor). Для индукции транскрипции Adh 1 GF14 должен сформировать комплекс с GBP, чтобы последний мог вступить в непосредственное взаимодействие с нуклеотидной последовательностью G-бокса.
Как отмечалось выше, биосинтез многих белков нормального метаболизма в бескислородных условиях ингибируется. Хотя гены этих белков при переходе к анаэробному метаболизму продолжают транскрибироваться с той же интенсивностью, которая наблюдается в аэробных условиях, трансляция их мРНК в значительной степени подавляется. Напротив, эффективность трансляции мРНК анаэробных генов, в частности Adh 1, при аноксии резко возрастает. Таким образом, регуляция экспрессии генов на уровне трансляции играет важную роль не только в подавлении синтеза белков нормального метаболизма, но и в индукции анаэробных белков.
8.5.5. В ПРОЦЕСС ОБРАЗОВАНИЯ АЭРЕНХИМЫ ПРИ ДЕФИЦИТЕ О2 ВОВЛЕЧЕН РАСТИТЕЛЬНЫЙ ГОРМОН ЭТИЛЕН
Синтез этилена в растениях стимулируют низкие концентрации кислорода в среде (0,003 — 0,012 МПа), т.е. условия гипоксии. Этот гормон индуцирует образование аэренхимы в коре корня. Аноксия по сравнению с гипоксией в меньшей степени способствует образованию аэренхимы, так как для синтеза этилена требуется кислород. Предшественник этилена — 1-аминоциклопропан-1-карбоновая кислота (АЦК). Количества АЦК-синтазы и АЦК-оксидазы (ферментов, вовлеченных в биосинтез этилена) существенным образом возрастают в ответ на гипоксию в корнях кукурузы. В пути передачи сигнала, который запускается этиленом и приводит к формированию аэренхимы, участвуют в качестве вторичного мессенжера ионы Са2+. Переданный сигнал индуцирует процессы, приводящие к гибели клеток в центральной части коры корня кукурузы и к их последующему лизису. Наиболее вероятно, что гибель клеток в этом случае — результат апоптоза, т. е. запрограммированной смерти. В лизисе клеток участвуют ферменты, осуществляющие деградацию клеточных стенок, — целлюлазы и ксилоглюканазы, синтезируемые в больших количествах в гипоксических корнях.
Этилен вовлечен также в ответ на затопление у глубоководных сортов риса. Такие сорта могут расти в воде, уровень которой достигает 4 м.
Затопление молодых растений приводит к усиленному образованию этилена, за которым следуют увеличение содержания в клетках АБК и возрастание отзывчивости растений на гибберелин. В результате скорость деления клеток в интеркалярной меристеме возрастает, а также усиливается растяжение клеток в междоузлиях. Благодаря этому соревнование в скорости между ростом и затоплением выигрывает рост растения, так что верхняя часть побега оказывается над поверхностью воды, снабжая кислородом затопленные части растения через аэренхиму.
Поддержание аэробного метаболизма в корнях при затоплении за счет формирования
О2 – проводящих путей, наряду с переходом на анаэробный метаболизм, можно рассматривать как важнейшую стратегию устойчивости растений к кислородному дефициту.








