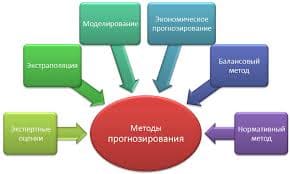
2018-01-21
2018-01-21 5794
5794Механизм электромеханического сопряжения в миокарде. У человека и млекопитающих структуры, которые отвечают за электромеханическое сопряжение в скелетных мышцах, в основном имеются и в волокнах сердца. Для миокарда характерна система поперечных трубочек (Т-система); особенно хорошо она развита в желудочках, где эти трубочки образуют продольные ответвления. Напротив, система продольных трубочек, служащих внутриклеточным резервуаром Са2+, в мышце сердца развита в меньшей степени, чем в скелетных мышцах. Как структурные, так и функциональные особенности миокарда свидетельствуют в пользу тесной взаимосвязи между внутриклеточными депо Са2+ и внеклеточной средой. Ключевым событием в сокращении служит вход в клетку Са2+ во время потенциала действия. Значение этого кальциевого тока состоит не только в том, что он увеличивает длительность потенциала действия и вследствие этого рефрактерного периода: перемещение кальция из наружной среды в клетку создает условия для регуляции силы сокращения. Однако количество кальция, входящего во время ПД, явно недостаточно для прямой активации сократительного аппарата; очевидно, большую роль играет выброс Са2+ из внутриклеточных депо, запускаемый входом Са2+ извне. Кроме того, входящие в клетку ионы пополняет запасы Са2+, обеспечивая последующие сокращения. Таким образом, потенциал действия влияет на сократимость по меньшей мере двумя путями. Он – играет роль пускового механизма («триггерное действие»), вызывающего сокращение путем высвобождения Са2+ (преимущественно из внутриклеточных депо); – обеспечивает пополнение внутриклеточных запасов Са2+ в фазе расслабления, необходимое для последующих сокращений. Механизмы регуляции сокращений. Целый ряд факторов оказывает косвенное влияние на сокращение миокарда, изменяя длительность потенциала действия и тем самым величину входящего тока Са2+. Примеры такого влияния — снижение силы сокращений вследствие укорочения ПД при повышении внеклеточной концентрации К+ или действии ацетилхолина и усиление сокращений в результате удлинения ПД при охлаждении. Увеличение частоты потенциалов действия влияет на сократимость так же, как и повышение их длительности (ритмоинотропная зависимость, усиление сокращений при нанесении парных стимулов, постэкстрасистолическаяпотенциация). Так называемый феномен лестницы (нарастание силы сокращений при их возобновлении после временной остановки) также связан с увеличением внутриклеточной фракции Са2+. Учитывая эти особенности сердечной мышцы, не приходится удивляться тому, что сила сокращений сердца быстро изменяется при изменении содержания Са2+ во внеклеточной жидкости. Удаление Са2+из внешней среды приводит к полному разобщению электромеханического сопряжения; потенциал действия при этом остается почти неизменным, но сокращений не происходит. Ряд веществ, блокирующих вход Са2+ во время потенциала действия, оказывает такой же эффект, как и удаление кальция из внешней среды. К таким веществам относятся так называемые антагонисты кальция (верапамил, нифедипин, дилтиазем) Напротив, при повышении внеклеточной концентрации Са2+ или при действии веществ, увеличивающих вход этого иона во время потенциала действия (адреналин, норадреналин), сократимость сердца увеличивается. В клинике для усиления сердечных сокращений используют так называемые сердечные гликозиды (препараты наперстянки, строфанта и т. д.). В соответствии с современными представлениями сердечные гликозиды повышают силу сокращений миокарда преимущественно путем подавления Nа+/К+-АТФазы (натриевого насоса), что приводит к повышению внутриклеточной концентрации Nа +. В результате снижается интенсивность обмена внутриклеточного Са2+ на внеклеточный Nа+, зависящего от трансмембранного градиента Nа, и Са2+ накапливается в клетке. Это дополнительное количество Са2+ запасается в депо и может быть использовано для активации сократительного аппарата.
Электрокардиография – метод регистрации электрической активности миокарда в ходе сердечного сокращения.
Регистрируется некоторый суммарный эффект активности клеток. На уровне отдельной клетки, при ее переходе в возбужденное состояние, на мембране происходит изменение электрического потенциала: на смену отрицательному потенциалу покоя приходит положительный потенциал действия. Этот процесс сопряжен с переносом ионов различного вида через мембрану. Клеточная мембрана в спокойном состоянии клетки поляризована таким образом: внутри клетки – минус, снаружи – плюс. При переходе клетки в возбужденное состояние ситуация на мембране меняется на противоположную: внутри клетки – плюс (преобладают положительные ионы), снаружи – минус. Эти изменения на клеточной мембране называются деполяризацией.
После того, как возбужденная клетка выполнила свою природную функцию, она возвращается в спокойное состояние; восстанавливаются и начальные концентрации ионов по обе стороны мембраны. Этот процесс называется реполяризацией.
Переходы клеток в возбужденное состояние и последующий их возврат в спокойное состояние носит массовый характер, и в различных участках миокарда начинается, происходит и заканчивается не одновременно. Поэтому можно говорить о волнах деполяризации и реполяризации, распространяющихся по сердцу в целом или по отдельным его частям - предсердиям, желудочкам, перегородкам.
Представлениям о сердце как органе, по которому распространяются волны деполяризации и реполяризации, хорошо соответствует модель, согласно которой сердце – это электрический диполь, электрическое поле которого ощутимо (благодаря проводящим тканям) далеко за пределами сердца, и может регистрироваться с помощью системы электродов, помещенных на поверхности тела.