 2018-03-09
2018-03-09 132
132
Студент должен разобраться в реакциях b-окисления и иметь представление об энергетической эффективности этого процесса. Так, при окислении одной молекулы пальмитиновой кислоты синтезируется 130 молекул АТФ. Необходимо задуматься над вопросами: на каких этапах окисления жирных кислот происходит образование АТФ, каков состав важнейших кислот?
Следует иметь в виду, что возможно также a- окисление (последовательное отщепление карбоксильного углерода в виде СО2) и w- окисление (конечный продукт - a, w-дикарбоновая кислота) жирных кислот.
Метаболизм кетоновых тел. При некоторых условиях (голодание, сахарный диабет, резкие смены диеты и т.д.) из ацетил-КоА в печени образуются кетоновые тела. Надо знать, что под этим термином подразумеваются: ацетоуксусная кислота, 3-гидроксимасляная кислота и ацетон. Механизм их образования из ацетил-КоА может быть представлен следующей схемой:

Если процессы образования кетоновых тел идут слишком интенсивно, в крови появляются ненормально высокие количества этих соединений, что приводит к понижению ее рН.
Биосинтез липидов. В живых организмах энергетический материал в больших количествах запасается в виде жирных кислот и триацилглицеринов.
Биосинтез жирных кислот. Необходимо запомнить, что биосинтез жирных кислот не является процессом, обратимым их окислению. Биосинтез жирных кислот имеет ряд характерных особенностей.
Основным местом синтеза жирных кислот, в отличие от окисления, является цитоплазма клеток. Исходным веществом для биосинтеза служит ацетил-КоА, который образуется из жирных кислот (главным образом), а также из пировиноградной кислоты и некоторых аминокислот (лейцин, изолейцин, лизин, тирозин) – здесь очень четко видна связь обмена углеводов, липидов и белков. В биосинтезе участвует также малонил-КоА, который можно назвать первым продуктом биосинтеза жирных кислот – (он образуется из ацетил-КоА и бикарбоната при участии АТФ и ацетил-КоА – карбоксилазы, содержащей биотин):

В качестве восстановителя в биосинтезе жирных кислот расходуется НАДФН, образующийся в пентозофосфатном цикле (50%) или при окислении малата:
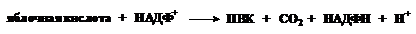
Конечным продуктом биосинтеза в цитоплазме является пальмитиновая кислота, а из нее уже получаются остальные насыщенные и ненасыщенные кислоты. Суммарную реакцию можно изобразить так:

Образующийся в митохондриях ацетил-КоА не способен переходить через мембрану в цитоплазме. Перенос его возможен двумя путями: с помощью цитрата (основной путь) и с помощью карнитина. В свою очередь, цитрат образуется в митохондриях из ацетил-КоА и оксалоацетата. После выхода в цитоплазму он вновь расщепляется АТФ-зависимым ферментом на исходные вещества:

Выше рассматривалась роль карнитина в переносе ацильных групп из цитоплазмы в митохондрии при окислении жирных кислот. По-видимому, он может выполнять эту роль и в обратном процессе, хотя для синтеза жирных кислот этот путь переноса ацильных групп не является главным.
Катализатором биосинтеза является мультиферментный комплекс, состоящий из семи ферментов – синтеза жирных кислот. Центральную роль в этой системе занимает ацилпереносящий белок (АПБ), с которым ковалентно связываются промежуточные продукты биосинтеза жирных кислот. Простетической группой в АПБ является 4-фосфопантотеин, к сульфгидрильной группе которого присоединяются ацильные остатки (подобно тому, как они присоединяются в КоАSH). Полагают, что 4-фосфопантотеин образует как бы «подвижную руку», переносящую остатки жирных кислот от активного центра одного фермента к активному центру другого. В синтезе содержится еще одна группа HS-, принадлежащая цистеину.
Для инициирования процесса биосинтеза жирных кислот требуется одна молекула ацетил-КоА: его углеродные атомы превращаются в конце концов в терминальную этильную группу пальмитиновой кислоты. Другие 7 двууглеродных фермента доставляются в форме малонил-КоА. Ацетильная группа переносится на SH-группу цистеинового остатка синтетазы, а малонильная группа – на SH-группу фосфопантотеина АПБ. Далее ацетильная и малонильная группы конденсируются с образованием ацетоацетильной группы. При этом ацетильная группа вытесняет свободную карбоксильную группу остатка малоната в виде СО2. Обратите внимание на то, что четырехуглеродная группа связана с SH-группой фосфопантотеина и остается здесь во время последующих реакций вплоть до образования бутирила. Далее бутирильная группа переносится на SH-группу цистеина, а с SH-группой фосфопантотеина связывается следующая малонильная группа. Бутирильная группа покидает SH-группу цистеина, замещает карбоксильную группу в малониле на HS-АПБ, и цикл повторяется.
Для уяснения всех стадий биосинтеза жирных кислот полезно обратиться к следующей схеме
Синтез триацилглицеринов. Свободные жирные кислоты в тканях и крови присутствуют в небольших количествах и в норме не накапливаются, а включаются в процесс синтеза жиров и фосфолипидов.
Синтез триацилглицеринов происходит, главным образом, в печени и жировой ткани из КоА-производных жирных кислот через промежуточное образование фосфатидной кислоты. Необходимо понять роль фосфатидной кислоты и запомнить последовательность реакций образования жиров.








