 2014-02-09
2014-02-09 1578
1578У ряда видов примитивных эукариот (Tetrahymena thermophila, Physarum polycephalum и др.) гены рРНК содержат особые нитроны (интроны группы 1), для которых характерен особый вид сплайсинга, осуществляемого без участия каких-либо белков и получившего название аутосплайсинг (сплайсинг типа I).
Коничев.
Процессинг тРНК эукариот.
Пре-тРНК, содержат вставку (интрон) вблизи антикодона. Вырезание этого интрона и лигирование (сшивание) остальной части молекулы-предшественника приводит к образованию зрелой тРНК (рис. 142). Необходимые для этого процесса нуклеазная (фосфодиэстеразная) лигазная и киназная реакции осуществляются, как полагают, одним полифункциональным ферментом.
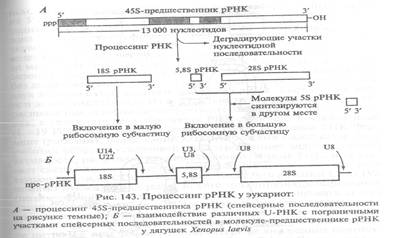
Процессинг рРНК высших организмов совершается в ядрах клеток на базе молекул-предшественников, транскрибируемых с генов, содержащихся в ДНК в виде тандемно повторяющихся копий (кластеров). В одном первичном транскрипте, имеющем у млекопитающих коэффициент седиментации 45S (-13 000 нуклеотидов), содержатся нуклеотидные последовательности 18S (-2000 нуклеотидов), 28S (-5000 нуклеотидов) и 5,8S (~ 160 нуклеотидов) рРНК, которые разделяются спейсерными последовательностями. Эндонуклеазное расщепление этого предшественника ведет к выщеплению зрелых рРНК (рис. 143).
5S рРНК человека, имеющая длину 120 нуклеотидов, кодируется и транскрибируется отдельно от других РНК с соответствующих генов, число которых в геноме достигает 2000. Синтез всех рРНК (кроме 5S рРНК) происходит в ядрышках с очень высокой скоростью. Тандемно повторяющиеся гены рРНК транскрибируются синхронно (рис. 144). Одновременная транскрипция генов рРНК и процессинг общего
транскрипта приводят к синхронному образованию пропорциональных количеств всех рРНК, необходимых для сборки рибосом.
Такие интроны встречаются также в генах рРНК митохондрий, хлоропластов, дрожжей и грибов, однако они не выявлены в генах позвоночных животных. Оказалось, что содержащаяся внутри 26S рРНК тетрахимены вставка (интрон) длиной 400 нуклеотидов способна сама осуществлять вырезание этого интрона и сшивание экзонов в присутствии Mg2+ и свободного гуанозина (либо его фосфорилированных производных). Таким образом была открыта аутокаталитическая функция РНК и положено начало изучению РНК-ферментов – рибозимов.
Схема аутосплайсинга рРНК у тетрахимены представлена на рис. 145_Коничев).
Альтернативный сплайсинг. (Коничев,с.295)
Некоторые интроны в молекуле пре-мРНК относятся к одному гену, но могут сплайсироваться разными способами. Это приводит к появлению разных по составу экзонов мРНК.
Такой альтернативный сплайсинг позволяет организму синтезировать разные по структуре и свойствам белки на базе одного гена. Он характерен для мРНК, при трансляции которых образуются родственные белки или изоформы, которые колируются одним геном.
Альтернативный сплайсинг. Несколько интронов, содержащихся в мРНК, могут сшиваться в разных комбинациях с образованием различных матричных последовательностей. Такая дифференциация путей созревания мРНК получила название альтернативного сплайсинга. Этот сплайсинг был впервые открыт у аденовирусов, у которых часть кодирующих последовательностей удаляется подобно интронам, а 5'-кэп соединяется с любым из других экзонов, и, таким образом, образуются различные мРНК, кодирующие разные белки (рис. 149). Этот вид сплайсинга позволяет небольшому числу первичных вирусных транскриптов кодировать значительное количество белков, в пропорциях, необходимых для существования вируса.
У эукариот альтернативный сплайсинг мРНК, содержащих большое количество интронов, создает возможность для возникновения изоформ белков (см. гл. 2), наборы которых могут существенно различаться в различных клетках, тканях и органах многоклеточных организмов
В формировании альтернативных мРНК могут быть задействованы три основные механизма.
Первый из них состоит в том, что для образования различных мРНК могут использоваться разные промоторы. В этом случае образуются транскрипты, имеющие разные по длине 5'-концы и разное количество экзонов. Такой механизм сплайсинга выявлен для пре-мРНК легкой цепи миозина позвоночных животных.
Второй тип альтернативного сплайсинга имеет место при изменении сайта полиаденилирования первичного транскрипта. В этом случае изменяются размеры и структура З'-концевого участка премРНК (рис. 150). Таким способом образуются два вида мРНК тяжелой цепи иммуноглобулинов.
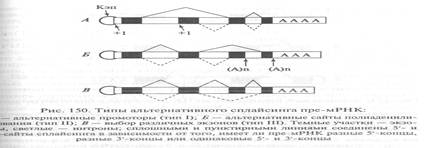
Третий тип альтернативного сплайсинга включает выбор различных экзонов из одинаковых пре-мРНК. При этом для формирования зрелых РНК могут использоваться различные экзоны, а часть из них не включается в сплайсинг. Таким образом происходит сплайсинг пре-мРНК тропонина Т скелетных мышц млекопитающих, содержащий 18 экзонов. При этом выбор экзонов зависит от стадии развития организма: экзон 16 присутствует в мРНК тропонина Т у взрослых, а экзон 17 — в мРНК этого белка у эмбрионов. Тонкие механизмы выбора сайта сплайсинга еще неизвестны. Однако одной из причин возникновения альтернативных продуктов могут служить мутации, приводящие к разрушению нормальных сайтов сплайсинга и возникновению новых сайтов (рис. 151). Такой механизм альтернативного сплайсинга выявлен у больных талассемией (группа наследственных заболеваний, при которых резко падает содержание гемоглобина). Потеря в результате мутации одного из сайтов сплайсинга еще не означает отмену сплайсинга в целом, так как оставшийся другой сайт «ищет» подходящий участок и может соединяться с таким скрытым, ранее не реализовавшимся сайтом сплайсинга с образованием новой мРНК. Таким образом, мутации в мозаичных генах эукариот могут стимулировать образование новых белков, а альтернативный сплайсинг может играть, решающую роль в эволюции высших организмов.
ПРОЦЕССИНГ У ПРОКАРИОТ (Коничев,с.283)
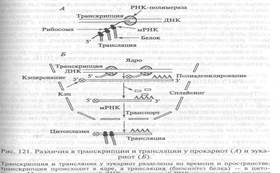
Процессинг у прокариот в основном затрагивает предшественники рРНК и тРНК, тогда как полицистроновые мРНК используются для трансляции (биосинтеза белка) сразу после их синтеза и даже еще до окончания транскрипции, так что биосинтез белка оказывается непосредственно сопряжен с транскрипцией (рис. 121).
Ранее считали, что полиаденилирование свойственно только мРНК эукариот. Однако в настоящее время установлено, что поли (А)-хвосты есть и у мРНК прокариот, но они короче, чем у мРНК эукариот (содержат от 14 до 60 остатков адениловой кислоты) и встречаются не у всех молекул мРНК. В отличие от эукариот полиаденилирование дестабилизирует структуру ряда мРНК бактерий.
Процессинг у бактерий необходим для образования зрелых молекул рРНК и тРНК. Первичный транскрипт, содержащий последовательности этих РНК, образуется на оперонах, включающих гены 16S рРНК, 23S рРНК, 5S рРНК и тРНК (рис. 141). Первичные транскрипты таких оперонов имеют длину 5000 нуклеотидов и помимо последовательностей рРНК содержат вставочные последовательности (спейсеры), существенные для процессинга.
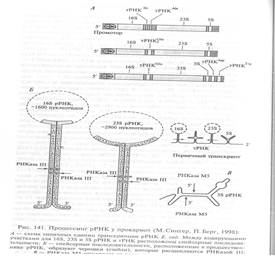
Вычленение индивидуальных рРНК происходит с участием эндонуклеаз, разрезающих первичные транскрипты в областях спейсеров.
Образующиеся после первичного эндонуклеотического расщепления пре-тРНК подвергаются дальнейшему процессингу при участии двух ферментов: эндонуклеазы Р и экзонуклеазы D (рис. 142). При этом пре-тРНК, содержащая более 100 нуклеотидных остатков, укорачивается с обоих концов до стандартного размера (70— 90 нуклеотидов). С 5'-конца на пре-тРНК действует эндонуклеаза Р (РНКазаР), а с З'-конца -экзонуклеаза D.
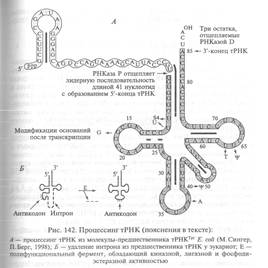
Эндонуклеазное действие РНКазы Р заключается в отщеплении 41 нуклеотида от 5'концевой (лидерной) последовательности предшественника и сразу приводит к формированию 5'-конца тРНК, представленного фосфорилированным гуанозином (5'-pG). Экзонуклеаза D, со своей стороны, отщепляет последовательно несколько нуклеотидов с З'-конца молекулы-предшественника, останавливаясь на аденозине, соседствующем с двумя цитидиловыми нуклеотидами, и таким образом формируется З'-конец (акцептирующий стебель) тРНК, содержащий стандартную последовательность ССА-ОН. Одновременно с процессингом рРНК и тРНК осуществляется модифицирование ряда азотистых оснований, приводящее, в частности, к образованию характерных для тРНК дигидроурацила (UH2), псевдоуридина (ψ) и тимина (Т). Ферменты, осуществляющие эти модификации, до сих пор не охарактеризованы. Эти модификации называются редактированием РНК (Рис. 142_Коничев Процессинг тРНК (пояснения в тексте)








