 2015-04-17
2015-04-17 12018
12018Основные физические характеристики звука - частота и интенсивность колебаний. Они и влияют на слуховое восприятие людей.
Периодом колебания называется время, в течение которого совершается одно полное колебание. Можно привести в пример качающийся маятник, когда он из крайнего левого положения перемещается в крайнее правое и возвращается обратно в исходное положение.
Частота колебаний - это число полных колебаний(периодов)за одну секунду. Эту единицу называют герцем (Гц). Чем больше частота колебаний, тем более высокий звук мы слышим, то есть звук имеет более высокий тон. В соответствии с принятой международной системой единиц, 1000 Гц называется килогерцем (кГц), а 1.000.000 - мегагерцем (МГц).
Распределение по частотам: слышимые звуки - в пределах 15Гц-20кГц, инфразвуки - ниже 15Гц; ультразвуки - в пределах 1,5104 - 109 Гц; гиперзвуки - в пределах 109 - 1013Гц.
Ухо человека наиболее чувствительно к звукам с частотой от 2000 до 5000 кГц. Наибольшая острота слуха наблюдается в возраст 15-20 лет. С возрастом слух ухудшается.
С периодом и частотой колебаний связано понятие о длине волны. Длиной звуковой волны называется расстояние между двумя последовательными сгущениями или разрежениями среды. На примере волн, распространяющихся на поверхности воды, - это расстояние между двумя гребнями.
Звуки различаются также по тембру. Основной тон звука сопровождается второстепенными тонами, которые всегда выше по частоте(обертона). Тембр - это качественная характеристика звука. Чем больше обертонов накладывается на основной тон, тем «сочнее» звук в музыкальном отношении.
Вторая основная характеристика - амплитуда колебаний. Это наибольшее отклонение от положения равновесия при гармонических колебаниях. На примере с маятником - максимальное отклонение его в крайнее левое положение, либо в крайнее правое положение. Амплитуда колебаний определяет интенсивность(силу) звука.
Сила звука, или его интенсивность, определяется количеством акустической энергии, протекающей за одну секунду через площадь в один квадратный сантиметр. Следовательно, интенсивность акустических волн зависит от величины акустического давления, создаваемого источником в среде.
С интенсивностью звука в свою очередь связана громкость. Чем больше интенсивность звука, тем он громче. Однако эти понятия не равнозначны. Громкость - это мера силы слухового ощущения, вызываемого звуком. Звук одинаковой интенсивности может создавать у различных людей неодинаковое по своей громкости слуховое восприятие. Каждый человек обладает своим порогом слышимости.
Звуки очень большой интенсивности человек перестаёт слышать и воспринимает их как ощущение давления и даже боли. Такую силу звука называют порогом болевого ощущения.
53. Путь звуковой волны. Звукопроведение. Звуковосприятие.
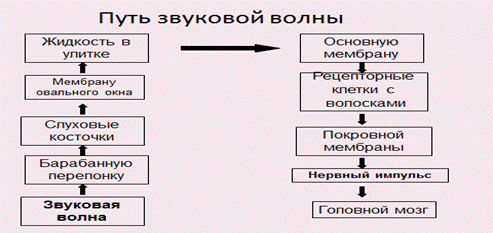
Функция звукопроведения состоит в передаче звуковых колебаний составными элементами наружного, среднего и внутреннего уха слуховым рецепторам.
В звукопроведении принимают участие ушная раковина, наружный слуховой проход, барабанная перепонка, слуховые косточки, кольцевая связка овального окна, вторичная барабанная перепонка, перилимфа, основная мембрана.
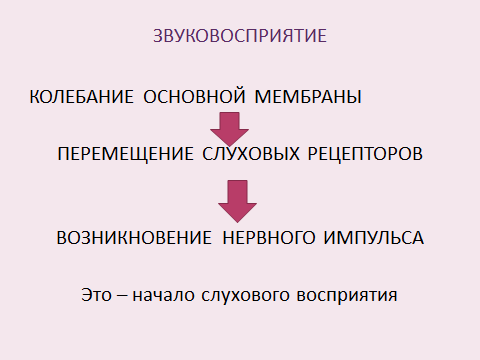
При раздражении волосковых клеток кортиева органа происходит превращение физической энергии звуковых колебаний в физиологический процесс нервного возбуждения. Это начало процесса слухового восприятия.
Область слухового восприятия 16-20000 Гц.
54. Область звукового восприятия. Чувствительность органа слуха.
ОБЛАСТЬ СЛУХОВОГО ВОСПРИЯТИЯ
16 – 20 000 Гц
Звуки с частотой ниже 16 Гц – инфразвуки
Звуки с частотой выше 20 000 Гц – ультразвуки
Периферический отдел слухового анализатора производит первичный анализ и преобразует физическую энергию звука в электрическую энергию нервного импульса. Проводящие пути передают импульс в мозговые центры. В коре головного мозга происходит превращение энергии нервного возбуждения в ощущение. Кора играет ведущую роль в работе слухового анализатора.
Ухо человека наиболее чувствительно к звукам от 500 до 4000 Гц – это речевой диапазон частот, (1000-3000 Гц).
Минимальная сила звука, способная вызвать ощущение едва слышимого звука – порог слышимости.
Чем ниже порог слышимости, тем выше чувствительность уха к данному звуку. При нормальном слухе величина порога слухового ощущения 0 дБ. При увеличении силы звука ощущение громкости звука усиливается, но при достижении определенной величины нарастание громкости прекращается и появляется ощущение боли – болевой порог. Расстояние между порогом слышимости и порогом неприятных ощущений в области средних частот – 130 дБ.
· Разностным порогом частоты называют минимальный прирост частоты звука к его первоначальной частоте – 3 Гц.
· Разностным порогом силы звука называют минимальный прирост силы звука, дающий усиление первоначальной громкости – 1 дБ.
Таким образом, область слухового восприятия у человека ограничена по высоте и силе звука.
55. Теории звукового восприятия.
Восприятие звуков различной высоты (частоты), согласно резонансной теории Гельмгольца,
обусловлено тем, что каждое волокно основной мембраны настроено на звук определенной частоты.
Так, звуки низкой частоты воспринимаются длинными волнами основной мембраны, расположенными
ближе к верхушке улитки, звуки высокой частоты воспринимаются короткими волокнами основной
мембраны, расположенными ближе к основанию улитки. При действии сложного звука возникают
колебания различных волокон мембраны.
В современной интерпретации резонансный механизм лежит в основе теории места, в соответствии
с которой в состояние колебания вступает вся мембрана. Однако максимальное отклонение основной
мембраны улитки происходит только в определенном месте. При увеличении частоты звуковых
колебаний максимальное отклонение основной мембраны смещается к основанию улитки, где
располагаются более короткие волокна основной мембраны, – у коротких волокон возможна более
высокая частота колебаний. Возбуждение волосковых клеток именно этого участка мембраны при
посредстве медиатора передается на волокна слухового нерва в виде определенного числа импульсов,
частота следования которых ниже частоты звуковых волн (лабильность нервных волокон не превышает
800 – 1000 Гц). Частота воспринимаемых звуковых волн достигает 20 000 Гц. Таким способом
осуществляется пространственный тип кодирования высоты и частоты звуковых сигналов.
При действии тонов примерно до 800 Гц кроме пространственного кодирования происходит еще и
временное (частотное) кодирование, при котором информация передается также по определенным
волокнам слухового нерва, но в виде импульсов (залпов), частота следования которых повторяет
частоту звуковых колебаний. Отдельные нейроны на разных уровнях слуховой сенсорной системы
настроены на определенную частоту звука, т.е. каждый нейрон имеет свой специфический частотный
порог, свою определенную частоту звука, на которую реакция нейрона максимальна. Таким образом,
каждый нейрон из всей совокупности звуков воспринимает лишь определенные достаточно узкие
участки частотного диапазона, не совпадающие между собой, а совокупности нейронов воспринимают
весь частотный диапазон слышимых звуков, что и обеспечивает полноценное слуховое восприятие.
Правомерность этого положения подтверждается результатами протезирования слуха человека, когда
электроды вживлялись в слуховой нерв, а его волокна раздражались электрическими импульсами
разных частот, которые соответствовали звукосочетаниям определенных слов и фраз, обеспечивая
смысловое восприятие речи.
Первая теория была создана британским физиком Резерфордом в 1886 году. Он предположил, что: а) звуковая волна заставляет вибрировать всю базилярную мембрану и частота вибраций соответствует частоте звука; б) частота вибраций мембраны задает частоту нервных импульсов, передаваемых по слуховому нерву. Так, тон частотой 1000 герц заставляет базилярную мембрану вибрировать 1000 раз в секунду, в результате чего волокна слухового нерва разряжаются с частотой 1000 импульсов в секунду, а мозг интерпретирует это как определенную высоту. Поскольку в этой теории предполагается, что высота зависит от изменений звука во времени, ее назвали временной теорией (ее называют также частотной теорией).
Гипотеза Резерфорда вскоре встретилась с серьезными проблемами. Было доказано, что нервные волокна могут передавать не более 1000 импульсов в секунду, и тогда неясно, как человек воспринимает высоту тона с частотой более 1000 герц. Вивер (Weaver, 1949) предложил способ спасения временной теории. Он предположил, что частоты выше 1000 герц кодируются различными группами нервных волокон, каждая из которых активируется в несколько разном темпе. Если, например, одна группа нейронов выдает 1000 импульсов в секунду, а затем 1 миллисекунду спустя другая группа нейронов начинает выдавать 1000 импульсов в секунду, то комбинация импульсов этих двух групп даст 2000 импульсов в секунду. Эту версию временной теории подкрепило открытие, что паттерн нервных импульсов в слуховом нерве повторяет форму волны стимульного тона, несмотря на то, что отдельные клетки реагируют не на каждое колебание (Rose et al., 1967).
Однако способность нервных волокон отслеживать форму волны обрывается примерно на частоте 4000 герц; тем не менее мы можем слышать высоту звука, содержащего гораздо более высокие частоты. Отсюда следует, что должно существовать другое средство кодирования высотного качества звука, по крайней мере на высоких частотах.
Другая теория восприятия высоты звука относится к 1683 году, когда французский анатом Жозеф Гишар Дювернье предположил, что частота кодируется высотой звука механически, путем резонанса (Green & Wier, 1984). Чтобы разобраться в этом предположении, полезно сначала рассмотреть пример резонанса. Когда ударяют по камертону, который находится рядом с пианино, струна пианино, настроенная на частоту камертона, начинает колебаться. Если мы говорим, что ухо работает по тому же принципу, это значит, что в нем есть некая структура, сходная по конструкции со струнным инструментом, причем различные ее части настроены на различные частоты, так что когда на ухо предъявляется некоторая частота, соответствующая часть этой структуры начинает колебаться. Эта идея была в общем правильной: такой структурой оказалась базилярная мембрана.
Как именно колеблется базилярная мембрана, не было известно до 1940 года, когда Георг фон Бекеши измерил ее движения при помощи маленьких отверстий, просверленных в улитках морских свинок и человеческих трупов. Учитывая результаты Бекеши, потребовалось модифицировать теорию локальности; базилярная мембрана вела себя не как пианино с раздельными струнами, а как простыня, которую встряхнули за один конец. В частности, Бекеши показал, что при большинстве частот вся базилярная мембрана приходит в движение, но место наиболее интенсивного движения зависит от конкретной частоты звучания. Высокие частоты вызывают вибрацию в ближнем конце базилярной мембраны; по мере повышения частоты паттерн вибрации сдвигается к овальному окошечку (Bekesy, 1960). За это и другие исследования слуха Бекеши получил в 1961 году Нобелевскую премию.
Как и временные теории, теория локальности объясняет многие, но не все явления восприятия высоты звука. Основные затруднения у теории локальности связаны с тонами низких частот. При частотах ниже 50 герц все части базилярной мембраны вибрируют примерно одинаково. Это значит, что все рецепторы активируются в равной степени, из чего следует, что у нас нет способа различения частот ниже 50 герц. На самом же деле мы можем различать частоту всего в 20 герц.
Таким образом, теории локальности затрудняются объяснить восприятие низкочастотных звуков, а временные теории — восприятие высоких частот. Все это навело на мысль, что восприятие высоты звука определяется как временными паттернами, так и паттернами локализации, причем временная теория объясняет восприятие низких частот, а теория локальности — восприятие высоких частот. Ясно, однако, что там, где один механизм отступает, начинает преобладать другой. На самом деле не исключено, что частоты от 1000 до 5000 герц обслуживаются обоими механизмами (Coren, Ward & Enns, 1999).
Поскольку наши уши и глаза играют столь важную роль в нашей повседневной жизни, были предприняты значительные усилия, направленные на то, чтобы заменить их на искусственные у индивидуумов, страдающих неизлечимыми дефектами этих органов. Некоторые из этих усилий описаны в рубрике «На переднем крае психологических исследований».
56. Стадии сна. Ритмы ЭЭГ при различных стадиях сна. Виды сна. Потребность во сне в различные периоды онтогенеза. Нарушения сна.
Общая характеристика. Сон – это особая активность мозга, при которой выключено сознание и
механизмы поддержания естественной позы, снижена чувствительность анализаторов. Засыпанию
способствует ряд факторов: соблюдение режима сна, т.е. сон в одно и то же время (циркадианный
биоритм), утомление нервных клеток, ослабление активности анализаторов (закрытие глаз, тишина),
удобная поза. Человек может спать и во время шума (шум от автомобилей на улице, невыключенное
радио и т.д.). Следует, однако, помнить, что шум отрицательно влияет на сон, нарушая его глубину,
последовательность фаз и тем самым ухудшая общее самочувствие. Поэтому спальню нужно, насколько
это возможно, изолировать от внешних раздражителей.
Признаки сна: 1) снижение уровня сознания; 2) зевание; 3) понижение чувствительности
анализаторов; 4) урежение сердцебиений и дыхания, снижение секреторной деятельности желез
(слюнных – сухость слизистой рта, слезных – жжение глаз, слипание век).
Продолжительность сна взрослых 7 – 8 ч в сутки. Однако известны случаи, когда люди длительное
время спали значительно меньше и сохраняли высокую работоспособность. Например, Наполеон I и Т.
Эдисон спали по 2 ч. К настоящему времени известно, что люди, спящие 7 – 8 ч в сутки, живут дольше
других при прочих равных условиях. Продолжительность сна у детей зависит от возраста.
Новорожденный спит около 20 ч в сутки, в возрасте 6 мес -15 ч. Естественная потребность во сне с
годами уменьшается. К концу первого года жизни продолжительность сна сокращается до 13 ч в сутки.
Средняя продолжительность сна у детей 2-го года – 12 ч, 9 лет – 10 ч, 13 – 15 лет – 9 ч, 16 – 19 лет – 8 ч
в сутки.
Структура сна. Весь период сна делится на две фазы: медленный и быстрый сон. Сонное состояние
мозга характеризуется возникновением в ЭЭГ «сонных веретен» (12 – 16 колебаний в 1 с) и
синхронизированными крупными медленными волнами ЭЭГ в -диапазоне. Такая фаза сна получила
название медленноволнового (ортодоксального) сна. Это состояние мозга периодически в течение ночи
заменяется быстрой низкоамплитудной десинхронизированной активностью (до 30 колебаний в 1 с),
которая напоминает ЭЭГ человека и животных во время бодрствования. Так как при этом сон не
прерывается, а по некоторым показателям становится даже более глубоким, то эта фаза сна в отличие от
предыдущей получила наименование парадоксального (быстрого) сна. Смена быстрого и медленного
сна происходит через равные промежутки времени со средней длительностью около 90 мин (один
цикл). При этом на медленный сон приходится около 80%, на быстрый – 20 % от всего периода сна.
Одной из характерных черт быстрого сна является возникновение быстрых движений глаз, более
сильное снижение тонуса мышц. На этом фоне у животных возникают различные движения: усов, ушей,
хвоста, подергивания лап, лизательные и сосательные движения, учащается и становится нерегулярным
дыхание, возникает неритмичный и частый пульс, повышается артериальное давление, усиливается
гормональная активность. Весьма существенно, что при этом активность мотонейронов спинного мозга
резко заторможена. В период медленного сна наблюдается урежение дыхания, пульса, снижение
артериального давления, общие движения туловища. Лишение животных парадоксального сна делает
их возбудимыми, раздражительными.
Рис. 9.2. Классификация стадий сна (А – Е) у человека с учетом особенностей ЭЭГ (по Лумису и др.;
Клайтману и др.). Три нижние кривые представляют собой одновременную запись ЭЭГ, ЭОГ и ЭМГ
указательного пальца во время БДГ-сна (со сновидениями). Обычно его эпизоды возникают в конце каждогоцикла сна
Для оценки глубины сна обычно используют электроэнцефалограмму (ЭЭГ). По особенностям ЭЭГ,
исходя из общепринятых стандартных критериев, выделяют четыре или пять стадий медленного сна. В
состоянии расслабленного бодрствования преобладает -ритм с изменчивой амплитудой (рис. 9.2). В
стадии А сна -ритм постепенно исчезает, между его эпизодами появляются все более длительные
интервалы с очень маленькими -волнами. Это соответствует переходу от бодрствования ко сну
(дремота), она длится несколько минут, причем некоторые авторы относят стадию А сна к
бодрствованию. Для стадии В сна (засыпание и самый поверхностный сон) характерны -волны. В
конце стадии над прецентральной областью мозга можно записать высокоамплитудные «вертекс-
зубцы» длительностью 3 – 5 с, предвещающие наступление стадии С сна (поверхностный сон). После
их появления спящий человек уже не различает слабые внешние раздражители. Характерной
особенностью биоэлектрической активности мозга в этой фазе служат веретенообразные всплески -
ритма («сонные веретена») и К-комплексы. В cmадии D сна (умеренно глубокий сон) регистрируются
быстрые -волны частотой 3,0–3,5 Гц, а в стадии Е сна (глубокий сон) - медленные
(синхронизированные) колебания, представляющие собой почти исключительно крайне медленные -
волны (частотой 0,7 – 1,2 Гц), на которые эпизодически накладываются мелкие -волны.
Рис. 9.3. Соотношение сна и бодрствования, а также БДГ-фазы и медленноволнового сна в различные периодыжизни человека. (по H.P. Roffward и др., 1966)
Наиболее существенное изменение в раннем возрасте – уменьшение обшей длительности сна и значительное снижение в нем доли БДГ-фазы
Затем развивается фаза быстрого сна, характеризующаяся десинхронизацией ЭЭГ (как в стадии В)
и эпизодами быстрых движений глаз (БДГ), которые можно наблюдать со стороны через сомкнутые
веки спящего или записывать методами электроокулографии (см. кривую ЭОГ на рис. 9.2).
Соотношение стадий быстрого и медленного сна и изменения их соотношения в онтогенезе
представлены на рис. 9.3. Остальная мускулатура в фазу быстрого сна, как и во время медленного сна,
атонична, за исключением возникающих иногда судорожных сокращений мышц лица или пальцев (см.
ЭМГ на рис. 9.2), сопровождающихся возрастанием частоты дыхания и сужением сосудов пальцев.
Сновидения – возникающие во сне образные представления и воспринимаемые как реальная
действительность. Детям и взрослым гораздо легче вспомнить содержание только что увиденного сна,
если их разбудить во время БДГ-фазы или тотчас после ее окончания; проснувшись в фазе
медленноволнового сна, человек часто не помнит сновидений. Отмечается высокая частота
воспоминаний в первом случае (60 – 90%) и существенно более низкая, причем значительно
колеблющаяся (от 1 до 74%), во втором. В то же время в медленноволновом сне наблюдается разговор,
снохождение и ночные страхи у детей. По некоторым данным, в 64 % пробуждений от медленного сна
человек рассказывает о психических переживаниях. Причем они, скорее, напоминают не сновидения, а
мысли, рассуждения. Между переживаниями во сне в медленном и парадоксальном сне существуют
значительные различия. В медленном сне во время сновидений зрительные картины менее четки, менее
аффективны, менее длительны и более реальны. Обнаружено, что даже когда людей или животных в
течение длительного времени лишали БДГ-сна, а следовательно, и сновидений, вопреки
существовавшим ранее предположениям, никаких продолжительных физических или психических
расстройств у них не возникало.
Факторы, побуждающие сновидения. 1. Предшествующая сну деятельность (дети продолжают
«играть» во сне, исследователь ставит эксперименты и т.д.). Например, известному физиологу О. Леви
приснилась модель опыта, с помощью которого он открыл медиаторный механизм передачи влияний с
симпатического и парасимпатического нервов на сердце. Менделееву сновидение помогло создать свою
знаменитую таблицу химических элементов. 2. Раздражители, действующие на организм во время сна.
Так, если приложить горячую грелку к ногам, спящему человеку может присниться сон, что он идет по
раскаленному песку. 3. Избыточная импульсация от переполненных или больных внутренних органов
может вызывать кошмарные сновидения. 4. Биологические потребности могут вызвать
соответствующие сновидения, например в случае отклонения показателей гомеостазиса.
Н.И. Касаткин (1973) полагает, что сновидения в период быстрого сна выполняют роль «стража»,
сигнализирующего о внутренних опасностях, ибо в сновидениях могут быть предсказаны заболевания
на 1 – 3 мес раньше их появления. Сны носят преимущественно зрительный характер. У
слепорожденных зрительные образы в снах отсутствуют и преобладают осязательные. К настоящему
времени установлено, что нет людей, не видящих сны, которые возникают в среднем 4 – 6 раз в ночь.
Если пробуждение наступает в стадии быстрого сна, то 70 – 90% людей детально и достаточно
эмоционально рассказывают о своих сновидениях, а если в медленном – лишь 7 – 10%. Часть
сновидений связана с сексуальной жизнью. Такой характер сновидений (у молодых и холостых людей
или при длительном половом воздержании) сопровождается поллюциями. В среднем 70% женщин
также видят сексуальные сны, в период которых может возникать оргазм. Сексуальные мотивы во сне
возникают у девушек в период менструаций.
57. Состояние бодрствования.
Бодрствование – состояние психики, характеризующееся достаточно высоким уровнем электрической активности мозга, свойственным активному взаимодействию индивида с внешним миром. Бодрствование является тем функциональным состоянием, на фоне которого разворачивается любая психическая деятельность. Значимость этого состояния для обеспечения эффективности деятельности при ее оптимальной физиологической стоимости чрезвычайно велика. Состояние бодрствования не является однородным. В нем выделяются активное бодрствование и спокойное бодрствование.
Одну из важнейших ролей в поддержании состояния бодрствования играет ретикулярная формация среднего мозга, от нейронов которой восходящие влияния идут к неспецифическим ядрам таламуса, а от них ко всем зонам коры больших полушарий. Бодрствование образует поле всевозможных сочетаний функций сознания – от состояния спокойного бодрствования через активное, напряженное бодрствование до выраженных аффектов.
В общих чертах схема нашей психики в состоянии бодрствования на основании данных объективной психологии выглядит следующим образом.
Природа раздражений, достигающих мозга, и вместе с тем восприятия имеют двойственный характер. Одни раздражения поступают в мозг от внутренних областей тела и обусловлены они разнообразными органическими процессами. Они возбуждают в мозгу различного рода органические впечатления, оставляющие в нем известные следы способные к оживлению.
Другой порядок раздражений проникает в мозг от воздействий, которые идут извне организма и влияют на мозг посредством, так называемых внешних воспринимающих органов. Они являются материальной основой внешних впечатлений, субъективным показателем которых служат ощущения. Часть внешних впечатлений и образуемых ими следов вступают в соотношение со сферой личности и становятся ее достоянием
Другие же внешние впечатления и их следы до поры, до времени оставаясь вне сферы личности, тем не менее, возбуждают те или другие внешние двигательные или иные реакции, которые в большинстве случаев не вступают в соотношение с личностью, — иначе говоря, остаются незамеченными нами. Сюда относится целый ряд психорефлекторных двигательных реакций, таких, как ходьба, мимические движения и множество других движений, которые принято считать автоматическими. Но с того момента, когда эти движения возбуждают реакцию сосредоточения, они уже вступают в соотношение со сферой личности и становятся в прямую от нее зависимость. Таким образом и бессознательная ассоциативная деятельность, вступая путем внутреннего сосредоточения в соотношение со сферой личности, становится как бы ее достоянием и становится от нее зависимой в том смысле, что может быть оживляема под влиянием личных потребностей
58. Механизмы регуляции сна и бодрствования.
Переход от бодрствования ко сну предполагает два возможных пути. Прежде всего не исключено,
что механизмы, поддерживающие состояние бодрствования, постепенно «утомляются». В соответствии
с такой точкой зрения сон - это пассивное явление, следствие снижения уровня бодрствования. Однако
не исключено и активное торможение механизмов, обеспечивающих бодрствование. И.П. Павлов
выделял два механизма развития сна, которые, по существу, подтверждают правомерность позиций
сторонников как пассивной, так и активной теории сна. С одной стороны, сон возникает как явления
охранительного торможения в результате сильного и длительного раздражения какого-либо
отдельного участка коры больших полушарий. С другой стороны, сон возникает как результат
внутреннего торможения, т.е. активного процесса формирования отрицательного условного
рефлекса. Важную роль в регуляции цикла сон – бодрствование играет ретикулярная формация ствола
мозга, где находится множество диффузно расположенных нейронов, аксоны которых идут почти ко
всем областям головного мозга, за исключением неокортекса. Роль РФ в цикле сон – бодрствование
была исследована в конце 1940-х годов учеными Г. Моруцци и Н. Мэгуном, обнаружившими, что
высокочастотное электрическое раздражение этой структуры у спящих кошек приводит к их
мгновенному пробуждению. И напротив, повреждения ретикулярной формации вызывают постоянный
сон, напоминающий кому; перерезка же только сенсорных трактов, проходящих через ствол мозга,
такого эффекта не дает. Наиболее ранними теориями сна были гуморальные. Фактор сна, лишенный
видовой специфичности, был выделен из ликвора коз, подвергнувшихся депривации сна. Согласно
сосудистой (циркуляторной или гемодинамической) теории сна, наступление сна связано со
снижением кровотока в мозге или с его усилением. Современные исследования показали, что в течение
сна действительно происходит колебание кровенаполнения мозга. Р. Лежандр и X. Пьерон (1910)
считали, что сон возникает в результате накопления токсических продуктов обмена вследствие
утомления (гипотоксины). Собакам долгое время не давали спать, а затем забивали, экстрагировали
вещества из мозга и вводили другим собакам. У последних развивались признаки крайнего утомления и
возникал глубокий сон. То же наблюдалось при «переносе» сыворотки крови или спинно-мозговой
жидкости.
В верхних отделах ствола мозга есть две области – ядра шва и голубое пятно, у нейронов которых
такие же обширные проекции, как и у нейронов ретикулярной формации, т.е. достигающие многих
областей ЦНС. Ядра шва захватывают срединную часть продолговатого мозга, моста и среднего мозга.
Разрушение их устраняет синхронизацию ЭЭГ и медленный сон. С помощью специальной методики
флуоресценции гистохимики показали, что нейроны ядер шва синтезируют серотонин и направляют
его через свои аксоны к ретикулярной формации, гипоталамусу, лимбической системе. Серотонин –
тормозной медиатор моноаминергической системы мозга. Блокада синтеза серотонина устраняет у
кошки медленный сон, у которой сохраняется лишь парадоксальный сон.
В среднем мозге (покрышка) обнаружено скопление нейронов, синтезирующих норадреналин
(голубое пятно). Стимуляция голубого пятна вызывает торможение нейронной активности во многих
структурах мозга при росте двигательного возбуждения животного и ЭЭГ-десинхронизации. Полагают,
что активирующее влияние голубого пятна осуществляется через механизм торможения тормозных
интернейронов. Ядра шва и голубое пятно действуют как антагонисты. Медиатором в клетках ядер
шва служит серотонин (5-гидрокситриптамин, 5-НТ), а голубого пятна – норадреналин. Разрушение
ядер шва у кошки приводит к полной бессоннице в течение нескольких дней; но за несколько
последующих недель сон нормализуется. Частичная бессонница может быть также вызвана
подавлением синтеза 5-НТ п-хлорфенилаланином. Ее можно устранить введением 5-
гидрокситриптофана, предшественника серотонина (последний не проникает через гематоэн-
цефалический барьер). Двустороннее разрушение голубого пятна приводит к полному исчезновению
БДГ-фаз, не влияя на медленноволновый сон. Истощение запасов серотонина и норадреналина под
влиянием резерпина вызывает, как и следовало ожидать, бессонницу. Однако оказалось, что нейроны
ядер шва наиболее активны и выделяют максимум серотонина не во время сна, а при бодрствовании.
Кроме того, возникновение БДГ, по-видимому, обусловлено активностью нейронов не столько голубого
пятна, сколько более диффузного подголубого ядра. Судя по результатам недавних экспериментов,
серотонин служит и медиатором в процессе пробуждения, и «гормоном сна» в бодрствующем
состоянии, стимулируя синтез или высвобождение «веществ сна» (факторов сна), которые в свою
очередь вызывают сон. Структуры таламуса выполняют функцию «пейсмекера» для вызова
ритмических потенциалов веретен во сне и -ритма в бодрствовании. Таламокортикальный механизм
можно рассматривать как механизм внутреннего торможения, способного изменять активность мозга
частично или глобально таким образом, что сенсорные, моторные и высшие функции мозга
подавляются.
Структуры, ответственные за медленный сон, находятся в каудальной части мозгового ствола,
главным образом – в продолговатом мозге. Наличие сходных гипногенных структур было установлено
также и в задней части моста. Двигательные и ЭЭГ-проявления фазы парадоксального сна связаны с
активацией структур в области моста. Эта фаза сна сокращается при эмоциональном стрессе, при этом
удлиняется период засыпания.
Рядом с голубым пятном имеется группа гигантских ретикулярных нейронов, которые направляют
свои аксоны вверх и вниз к различным структурам мозга. В бодрствовании и медленном сне эти
нейроны малоактивны, но их активность весьма высока во время парадоксального сна.
Были сделаны попытки обнаружить особые вещества либо после длительного лишения сна, либо у
спящего человека. Первый из этих подходов основан на предположении о том, что фактор(ы) сна во
время бодрствования накапливаются до вызывающего сон уровня, а второй – на гипотезе, согласно
которой они образуются или выделяются во сне.
Оба подхода дали определенные результаты. Так, при проверке первой гипотезы из мочи и
спинномозговой жидкости человека и животных был выделен небольшой глюкопептид – фактор S,
вызывающий медленноволновый сон при введении другим животным. Существует, по-видимому, и
фактор сна с БДГ. Второй подход привел к открытию индуцирующего глубокий сон нонапептида (в
настоящее время он уже синтезирован), так называемого пептида -сна (SIP, delta-sleep inducing
peptide). Однако пока неизвестно, играют ли эти и многие другие «вещества сна», обнаруженные при
проверке обеих гипотез, какую-либо роль в его физиологической регуляции. Более того, выделенные
пептиды часто вызывают сон лишь у животных определенного вида; кроме того, он возникает и под
действием других веществ.
Однако сросшиеся девочки-близнецы могли спать порознь, что свидетельствует о второстепенной
роли гуморальных факторов и решающей роли в развитии сна нервной системы.
Развивается представление о том, что цикл бодрствование – сон обеспечивается системой двух
центров. К. Экономо на основе клинических наблюдений больных с повреждениями различных
участков гипоталамуса предположил, что центр бодрствования локализован в заднем, а центр сна – в
его передних отделах. С. Рэнсон, производя локальные повреждения различных участков гипоталамуса,
подтвердил это мнение. В настоящее время считают, что гипоталамус является критической зоной для
регулирования цикла бодрствование – сон. Это мнение подтверждается и тем, что как высокочастотное,
так и низкочастотное электрическое раздражение преоптической области гипоталамуса вызывает
синхронизацию электроэнцефалограммы и поведенческий сон. Противоположный эффект, а именно
поведенческое и электроэнцефалографическое пробуждение Т.Н. Ониани наблюдал при раздражении
заднего гипоталамуса. Это позволяет предположить наличие реципрокного взаимоотношения между
передней и задней областями гипоталамуса и его значение для регуляции чередования различных фаз
цикла бодрствование – сон. По данным Т.Н. Ониани, в цикле бодрствование – сон мультинейронная
активность ретикулярной формации.

