 2015-04-30
2015-04-30 1572
15721. Способность к образованию внутренних солей (биполярных, или «цвиттер» – ионов). Карбоксильная группа проявляет кислотные свойства, диссоциирует с отщеплением протона. Амино-группа проявляет основные свойства – способна протонироваться по неподеленной электронной паре атома азота. В результате в водных растворах и в кристаллическом состоянии аминокислоты существуют в виде внутренних солей:
 +
+

 NH2-CH-COOH NH3-CH-COO-
NH2-CH-COOH NH3-CH-COO-
½ ® ½
R R
Суммарный заряд молекулы зависит от строения радикала R и рН среды:
а) моноаминомонокарбоновая кислота:
OH- + H+ +
[NH2-CH2-COO-] -1 ® [NH3-CH2-COO -]0 ®[ NH3-CH2-COOH]+1
б) моноаминодикарбоновая кислота:

 + +
+ +
NH2-CH-COO- -2 NH3-CH-COO- -1 NH3-CH-COO- °
½ HO- ½ Н+ ½
CH2 ® CH2 ® CH2
½ ½ ½
COO- COO- COOН


2Н+

 +
+
NH3-CH-COOH +1
½
CH2
½
СOOH
в) диаминомонокарбоновая кислота:
 | |||||||||||
 |  | |  | ||||||||
| |||||||||||
NH2-CH-COO- ° + NH3-CH-COO- +1 + NH3-CH-COOH +2
½ HO- ½ Н+ ½
(CH2 )4 ® (CH2)4 ® (CH2)4 - ½ ½ ½
+ NH3 +NH3 +NH3
¯2НО-


NH2-CH-COO- -1
½
(CH2)4
½
NH2
Свойство аминокислот изменять заряд при определенном значении рН, а, следовательно, двигаться в электрическом поле к разным электродам лежит в основе метода электрофореза, который используется для разделения смесей аминокислот и белков.
Значение рН среды, при котором суммарный заряд молекулы равен нулю, называется изоэлектрической точкой (ИЭТ, или рНi). В ИЭТ молекула теряет электрофоретическую подвижность, снижается растворимость белка в воде, он может выпадать в осадок.
2. Способность к образованию комплексных солей с ионами тяжелых металлов:
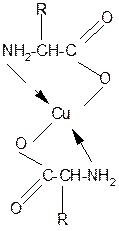 |
··
 2NH2-CH-C=O + Cu2+ +2Н+
2NH2-CH-C=O + Cu2+ +2Н+
½ ½
R OH
··
Комплексная соль Cu2+ хорошо растворима в воде и окрашена в ярко-синий цвет, что может быть использовано как качественная реакция на a- аминокислоты.
3. Реакция декарбоксилирования:
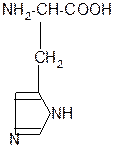
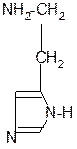 in vivo
in vivo


декарбоксилаза
CO2 +
 in vitro
in vitro
 |
t°, Ba(OH)2
гистидин гистамин
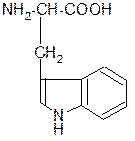
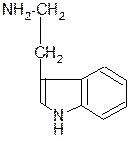
 СО2 +
СО2 +
триптофан триптамин
В результате реакции декарбоксилирования a- аминокислот образуются биогенные амины, обладающие сильной физиологической активностью.
4. Реакции дезаминирования:
а) дезаминирование по Ван-Слайку in vitro:
 NH2-CH2-COOH + HONO N2 + HO-CH2-COOH + H2O
NH2-CH2-COOH + HONO N2 + HO-CH2-COOH + H2O
глицин гликолевая кислота
По объему выделяющегося газообразного азота судят о количестве аминокислоты в растворе.
б) неокислительное дезаминирование in vivo протекает у низших организмов (бактерий, грибов):
-NH3
 HOOCCH 2 ®CH®COOH HOOC –CH=CH-COOH
HOOCCH 2 ®CH®COOH HOOC –CH=CH-COOH
¯ фумаровая кислота
NH2
аспарагиновая кислота
в) окислительное дезаминирование in vivo:
фермент, НАД+



 HOOC – (CH2)2 – C H _ COOH HOOC-(CH2)2-C-COOH
HOOC – (CH2)2 – C H _ COOH HOOC-(CH2)2-C-COOH
½ -НАД.H;-H+
NH 2 NH
глутаминовая кислота
H2O
 HOOC-(CH2)2-C-COOH
HOOC-(CH2)2-C-COOH

 -NH3
-NH3
O
a- кетоглутаровая кислота
Реакция протекает под действием фермента глутаматдегидрогеназы и кофермента НАД+.
5. Образование оснований Шиффа:
 H-C=O + H2N-CH2-COOH H2C-N H -CH2-COOH H2O +
H-C=O + H2N-CH2-COOH H2C-N H -CH2-COOH H2O +
½ глицин ½
H OH
+ CH2=N-CH2-COOH
метилениминокислота
Реакция лежит в основе метода формольного титрования по Зеренсену. Сами аминокислоты вследствие амфотерности не могут быть оттитрованы щелочью. Иминокислота содержит свободную карбоксильную группу и может быть определена количественно титрованием щелочью.
6. Взаимодействие с 2,4-динитрофторбензолом – ДНФБ (реактивом Сенджера):
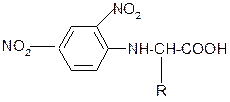 | |||
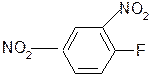 | |||
 + H2N-CH-COOH
+ H2N-CH-COOH
½ –HF
R
ДНФБ a-аминокислота ДНБ-производное
ДНБ-производное аминокислоты может быть выделено и идентифицировано хроматографически. Метод служит для определения аминокислотной последовательности белка.
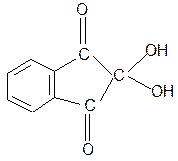 7. Нингидриновая реакция обусловлена наличием a- аминогруппы, качественная реакция на a- аминокислоты, пептиды и белки:
7. Нингидриновая реакция обусловлена наличием a- аминогруппы, качественная реакция на a- аминокислоты, пептиды и белки:
t°
 + NH2-CH-COOH
+ NH2-CH-COOH
2 ½
R
a- аминокислота
нингидрин
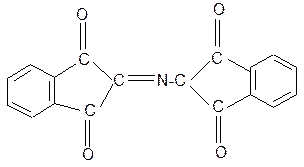 |
 H + CO2 + 3H2O + R-CHO
H + CO2 + 3H2O + R-CHO
продукт сине-фиолетового цвета
8. Реакция трансаминирования (переаминирования) in vivo:
фермент трансфераза

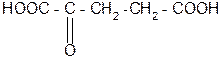 NH2-CH-COOH +
NH2-CH-COOH +
½ пиридоксальфосфат
CH3 (вит. В6)
аланин a- кетоглутаровая
кислота
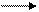 О=С-СOOH + НООС-СH-(СH2)2-СOOH
О=С-СOOH + НООС-СH-(СH2)2-СOOH
½ ½
CH3 NH2
пировиноградная глутаминовая
кислота кислота
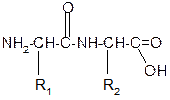 9. Способность к образованию полипептидов. Карбоксильная группа одной аминокислоты может реагировать с аминогруппой другой аминокислоты с образованием пептидной связи:
9. Способность к образованию полипептидов. Карбоксильная группа одной аминокислоты может реагировать с аминогруппой другой аминокислоты с образованием пептидной связи:
-H2O
 NH2-CH-C=O + H -NH-CH-C=O
NH2-CH-C=O + H -NH-CH-C=O
½ ½ ½ ½
R 1 OH R2 OH
Дипептид
Последовательность аминокислот, соединенных между собой пептидными связями, является первичной структурой белков.
Процесс, обратный образованию пептида, называется гидролизом.
Существуют три вида гидролиза белка: кислотный, щелочной и ферментативный. Результатом является образование смеси аминокислот, которые могут быть разделены и идентифицированы методами хроматографии или электрофореза. Таким образом определяют аминокислотный состав белка.
Рассмотрим пример:
Составить трипептид Глу-Асн-Про и дать ему полную характеристику.
 Решение:
Решение:
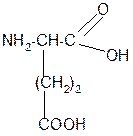 -2H2O
-2H2O

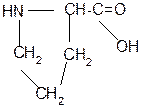 + +
+ +
 |
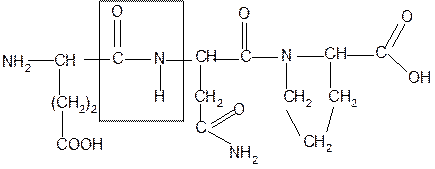 | |||
 | |||
глутамил-аспарагинил-пролин
В названии пептида окончания аминокислот меняются на «ил», кроме последней аминокислоты.
Характеристика пептида. 1. Реакция пептида на индикатор – кислотная, так как в пептиде две кислотные группы – СOOH и одна основная –NH2
(группа – С=О не проявляет основных свойств – см. классификацию)
½
NH2
2. Суммарный заряд пептида в водной среде:


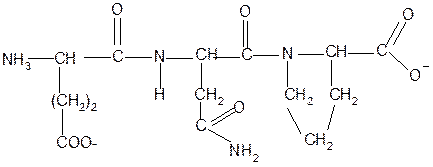 -1
-1
+

 3. Изменение суммарного заряда с изменением рН среды. Запишем пептид в упрощенном виде, выделив лишь заряженные группы и обозначив остальную часть пептида радикалом R: +
3. Изменение суммарного заряда с изменением рН среды. Запишем пептид в упрощенном виде, выделив лишь заряженные группы и обозначив остальную часть пептида радикалом R: +

 0 +1
0 +1 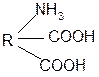

 -1 H+ + H+
-1 H+ + H+ 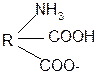


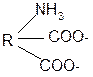 +
+
 OH-
OH-
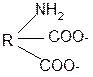

 -2
-2
4. Изоэлектрическая точка пептида (определение см. выше) лежит в слабокислой среде.
5. Поверхностные свойства пептида зависят от соотношения гидрофильных и гидрофобных боковых радикалов и концевых групп.
В пептиде содержатся 4 гидрофильные группы: 2-СOOH, -C=O, +NH3
и одна гидрофобная: радикал пролина ½
NH2
Гидрофильных групп больше, поэтому поверхность пептида гидрофильна, он растворим в воде.
Пептиды являются продуктом частичного гидролиза белков. Но многие пептиды присутствуют в свободном состоянии в клетках и тканях и выполняют специфические биологические функции. К ним относятся гормоны, антибиотики и другие соединения, обладающие высокой биологической активностью.
Самостоятельное значение пептидов в процессах жизнедеятельности человеческого организма велико.
В нервной ткани выделены нейропептиды, влияющие на функции нервной системы: энкефалины, эндорфины – «опиоидные» пептиды, аналогично морфину подавляющие боль; пептиды, действующие на сон; пептиды памяти и др.
Известны гормоны пептидной природы: окситоцин, вазопрессин (гормоны задней доли гипофиза), меланоцитстимулирующий гормон (выделяется средней долей гипофиза), адренокортикотропный гормон (АКТГ) (передней доли гипофиза), глюкагон (поджелудочной железы), гормоны желудочно-кишечного тракта и др.
К пептидам относятся токсины, выделенные из бледной поганки, токсины яда пчел, змей, скорпионов, морских позвоночных.
В медицине используются пептиды-антибиотики: грамицидин S, актиномицин и др.; пептиды-регуляторы иммунитета: тафцин; пептиды – заменители сахара: аспартам и т.д.
Строение пептидной связи
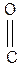

 ..
..
¾ ¾ N ¾
½
H
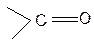 Атом углерода пептидной связи находится в sp2 –гибридизации. Неподеленная электронная пара атома азота вступает в сопряжение с p- связью. Таким образом, пептидная связь представляет собой трехцентровую r,p- сопряженную систему.
Атом углерода пептидной связи находится в sp2 –гибридизации. Неподеленная электронная пара атома азота вступает в сопряжение с p- связью. Таким образом, пептидная связь представляет собой трехцентровую r,p- сопряженную систему.
.
.
:…. ….
¾ ¾ N ¾
½
H
В результате связь C-N приобретает характер двоесвязанности, становится «полуторной». Вращение вокруг этой связи затруднено. В большинстве природных белков и пептидов имеет место транс-конфигурация пептидной связи, что важно для стабилизации вторичной структуры.
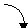
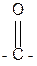 Для пептидной связи характерно явление лактам-лактимной таутомерии:
Для пептидной связи характерно явление лактам-лактимной таутомерии:
 OH
OH
½

 N ¾ ¾ C = N ¾
N ¾ ¾ C = N ¾
½
H
лактам лактим
За счет лактимной формы пептиды дают качественную цветную реакцию с ионами Сu2+ аналогично биурету – веществу, получаемому из 2 молекул мочевины:
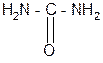

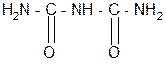 +
+
-NH3
мочевина биурет
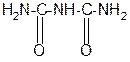

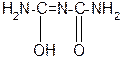

лактамная форма биурета лактимная форма биурета
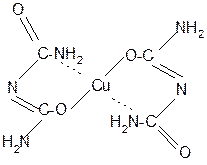 |
Эта реакция называется биуретовой. Она используется как качественная реакция для обнаружения пептидов и белков.
Тема 8. Углеводы
Цель занятия: закрепить и расширить знания принципов стереоизомерии, таутомерного равновесия и химических свойств, важнейших моно-, ди- и полисахаридов, участвующих в процессах жизнедеятельности, а также приобрести практические навыки идентификации углеводов.

