 2015-05-10
2015-05-10 1349
1349Нуклеиновые кислоты - сложные полимеры биологического происхождения. Они разделяются на две группы: дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые кислоты (РНК). Различие между этими группами кислот заключается, в том что в состав ДНК входит в качестве фрагментов полимерной цепи 2-D-дезоксирибоза, а в РНК в качестве фрагментов цепи входит D-рибоза. В качестве фрагментов макромолекулы, являющихся ответвлениями от полимерной цепи выступают пуриновые и пиримидино-вые основания. В ДНК это аденин, гуанин, цитозин и тимин, а в РНК аденин, гуанин, ци-тозин и урацил. Следует отметить, что в состав ДНК микроорганизмов входят и другие азотистые основания, впрочем не сильно отличающиеся (на группы СН3- и -СН2ОН).
Полимерная цепь ДНК построена следующим образом: за счет отрыва протона от гидроксилов в положениях 5' и 3' дезоксирибозы и гидроксилов от фосфорной кислоты с выделением воды (т.е. это поликонденсация) образуется полимерная цепь. Азотистые основания (АО) в образовании полимерной цепи не участвуют. Ниже представлена схема образования фрагмента макромолекулы ДНК из нуклеотидов:
Прямоугольником выделено многократно повторяющееся элементарное звено, которым, как видно, является остаток нуклеотида. В нём в качестве азотистого основания может быть один из четырех остатков: аденина, гуанина, цитозина или тимина.
РНК построена аналогично. Ниже приведен участок полимерной цепи РНК. Элементарным звеном в этой цепи, то есть многократно повторяющимся участком является нуклеотид, включающий в себя остатки ортофосфорной кислоты, рибофуранозы и одного из следующих оснований: аденина, гуанина, цитозина или урацила:
Когда из биологических тканей были выделены нуклеиновые кислоты и проведены работы по изучению состава и строения нуклеиновых кислот, то обнаружились следующие закономерности:
1) Молярное содержание аденина равно молярному содержанию тимина.
2) Молярное содержание гуанина равно молярному содержанию цитозина.
3) Суммарное молярное содержание пуриновых нуклеотидов равно суммарному со
держанию пиримидиновых нуклеотидов.
4) Суммарное молярное содержание нуклеотидов, имеющих в положении 6 и 4 амино
группу, равно суммарному молярному содержанию нуклеотидов, имеющих в поло
жении 6 и 4 карбонильную группу.
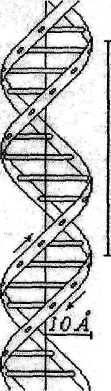
С учётом установленных закономерностей и на основании данных рентгено-структурного анализа и спектральных данных Уотсон и Крик в 1953 году создали модель пространственной конфигурации ДНК. Эта модель совмещает простоту и удивительную стройность и до самого последнего времени согласуется с новыми экспериментальными данными. Согласно модели Уотсона и Крика молекула ДНК состоит из двух полинуклео-тидных цепей, имеющих в качестве первичной структуры цепь из чередующихся фрагментов молекул 2-D-дезоксирибозы и фосфорной кислоты. Две такие цепи обвиваются вокруг общего цилиндра с одинаковым шагом винтовой линии.
При этом одна ветвь отстаёт от другой на полшага. Получается как бы «двухзаходная резьба» (как на импортных шурупах). Гетероциклические основания при этом обращены внутрь цилиндра и могут поворачиваться вокруг C-N гликозидной связи (а возможно и вокруг связи О-С3 и О-С3' дезоксирибозного фрагмента) так, что азотистые основания оказываются в одной плоскости. Упомянутые выше экспериментальные данные (пункты а-г) позволили Уотсону и Крику предположить, что в одну плоскость могут попасть только соответствующие друг другу по расположению амино- и карбонильных групп пуриновое с одной стороны и пиримидиновое с другой стороны основания. Такие основания называются комплементарными, то есть взаимно дополняющими. Нетрудно догадаться, что каждый гуанин должен быть комплементарен цитозину, так как будучи в одной плоскости и расположившись определённым образом они образуют три водородных связи:
Аналогично для пары - аденин-тимин, но образуются только две водородные связи:
Получается две цепи сцепленные «тяжами» из азотистых оснований, связанных довольно прочно (Г-Ц=60-80 кДж/моль; А-Т=40-56 кДж/моль)
Небольшой фрагмент ДНК можно схематически изобразить так:
Фрагмент ДНК часто изображают в виде двух спиралей скреплённых тяжами из комплементарных азотистых оснований. Расстояние между плоскостями двух соседних слоев, состоящих из оснований составляет 3,4 Ả, что очень близко к расстоянию между слоями в графите - 3,35 Ả. Один виток - 34 Ả, в нем уложено десять слоев из комплементарных азотистых оснований. Угол поворота между линиями, соединяющими места прикрепления азотистых оснований к дезоксирибозе (атомы С3 рибозы одной и другой ветви) составляет 360°. Диаметр цилиндра около 20 Ả. Считается, что 11 Ả из них занимает
| 34А |
 плоская система комплементарной пары А-Т или Г-Ц. Остальные
плоская система комплементарной пары А-Т или Г-Ц. Остальные
4,5 (20-11=9) Ả, по-видимому занимают остатки дезоксирибозы и
фосфорной кислоты, расположенные под углом как к оси цилиндра,
так и к воображаемым линиям, соединяющим атомы углерода (С1) в
остатках дезоксирибозы с двух сторон от комплементарных азотистых оснований. '
| 3,4А |
| Схема, двух- спиральной структуры ДНК (модель Уотсона и Крика Крика). |
Первый угол обеспечивает подъём по винтовой линии, а второй уменьшает размеры цилиндра. Не будь этого угла поворота, то есть, если бы оба фрагмента дезоксирибозы (их размер ~3 Ả) лежали в той же плоскости, что и азотистые основания, то диаметр цилиндра был бы на много больше: (3+5) *2=1б Ả 16+11=27 Ả. Тот и другой углы обеспечиваются валентным углом между плоскостью гетероцикла и плоскостью фуранозного кольца дезоксирибозы, а также поворотом этой плоскости вокруг связи N3(9)-C.
Число нуклеотидньгх единиц в цепи ДНК составляет от 3 000 до 10 000 000.Это означает, что молекулярная масса составит (а МА+ а Мт + в Мг + в Мц) 2n, где n - число нуклеотидных единиц, МА, Мт, MГ и МЦ - молярные массы мононуклеотидов из аденина, тимина, гуанина и цитозина соответственно. Коофициенты а и b - доли (от единицы) тех или иных нуклеотидов. Доля заметно колеблется для различных ДНК. Так, например, соотношение (Г+Ц) / (А+Т) колеблется для разных видов, (см. Таблицу 1)
Молярная масса большинства известных ДНК составляет от 4 до 16 миллионов, а для некоторых фагов 150 миллионов г/моль.
ДНК хорошо растворяется в воде, в слабых и даже крепких солевых растворах, образуя вязкую бесцветную жидкость. Она осаждается 2-3 объёмами 96% этанола, образуя студнеобразный или волокнистый осадок. ДНК денатурируется при нагревании водных растворов до 100°С, при подкислении до рН = 1-2 или подщелачивании выше рН = 10-12. При кислотном гидролизе от ДНК отщепляются пуриновые основания: аденин и гуанин. При нагревании с гидразином, наоборот, разрушаются пиримидиновые основания: цитозин и тимин. При действии на ДНК фермента дезоксирибонуклеазы происходит гидролиз фосфорноэфирных связей ДНК и она разбивается на нуклеотиды (моно). При помоши ферментов поджелудочной железы удаётся получить смесь тетра-, три- и динук Роль ДНК в организме состоит (по меньшей мере) в следующем. С помощью определённого кода на каждой из ветвей ДНК записывается вся наследственная информация. Эта информация потом считывается с помощью построения РНК комплементарной, участкам каждой ветви ДНК. На РНК как на матрице строится белок. Установлено, например, что сочетанию в цепи РНК трёх последовательно расположенных урацилов в белке соответствует фенилаланин. То есть, если взять раствор смеси аминокислот и поместить в этот же сосуд полинуклеотид, состоящий только из уридинмонофосфатов, то быстро при комнатной температуре синтезируется белок, состоящий только из фенилала- нина, а все остальные аминокислоты, останутся в растворе свободными даже если их концентрации были больше, чем концентрация фенилаланина у каждой. Таким образом нуклеиновые кислоты абсолютно селективные катализаторы поликонденсации аминокислот. РНК является той матрицей, на которой идёт синтез белка.
ДНК в клетках живых организмов не распределена равномерно (диффузно), а организована в особые структуры, связанные с передачей наследственных свойств. Эти структуры составляют основу так называемого ядерного вещества клеток - хроматина. Они представлены в виде хромосом и у высших организмов локализованы в клеточном ядре, а у большинства бактерий и других низших организмов в протоплазме, в виде нук-леидов. Хромосома представляет собой единый комплекс, состоящий из ДНК и молекул белка.
Таблица 1
| Вид | Показатели специфичности | |||||||||||||||||
| В ДНК: Г + Ц / А + Т | В РНК: Г + Ц / А + У | |||||||||||||||||
| Животные: | ||||||||||||||||||
Высшие растения:
| ||||||||||||||||||
| Грибы: Шампиньон Аспергилл | 0,80 1,00 | 1,01 1,23 | ||||||||||||||||
| Бактерии:Туберкулёзная палочка Тифозная бактерия Стафилококк | 2,08 1,13 0,53 | 1,45 1,21 1,05 | ||||||||||||||||
| Вирусы: Табачной мозаики Желтой мозаики репы | 0,78 1,23 |
Генетическая роль ДНК (впервые доказана экспериментально ещё в 1944 году) связана с двумя процессами:
1) точным воспроизведением самой себя, то есть способностью к редупликации
(самоудвоению). Это необходимо для того, чтобы у потомков была точно такая же
ДНК, как у родительских клеток;
2) определяющим влиянием на обмен веществ в клетке и, следовательно, на её биоло-
гические свойства. Это влияние ДНК не является прямым и непосредственным, а
идёт различными путями через белковый синтез.

