 2015-06-16
2015-06-16 1042
1042Рекомендовано Комитетом по высшей школе Миннауки России в качестве учебника для студентов биологических специальностей университетов
ИЗДАТЕЛЬСТВО МОСКОВСКОГО УНИВЕРСИТЕТА 1992
I. Вводная часть
o Глава 1. Исторический очерк (30К)
o Глава 2. Положение микроогранизмов в системе животного мира
o Глава 3. Размеры микроорганизмов
II. Мир прокариот
o Глава 4. Строение прокариотной клетки (103К)
o Глава 5. Морфологическая дифференцировка и уровни клеточной организации прокариот (38К)
o Глава 6. Общая характеристика конструктивного метаболизма прокариот (33К)
o Глава 7. Энергетический метаболизм прокариот (60К)
o Глава 8. Регуляторные системы у прокариот (34К)
o Глава 9. Прокариоты и факторы внешней среды (41К)
o Глава 10. Генетические механизмы эволюции прокариот (37К)
o Глава 11. Систематика прокариот. Группы прокариотных организмов (78К)
o Глава 12. Проблемы происхождения и эволюции жизни. Возникновение первичной клетки (54К)
III. Эволюция энергетических процессов у эубактерий (134К)
o Глава 13. Брожение. Типы жизни, основанные на субстратном фосфорилировании (134К)
o Глава 14. Фотосинтез. Типы жизни, основанные на фотофосфорилировании (156К)
o Глава 15. Молекулярный кислород как фактор эволюции (80К)
o Глава 16. Дыхание. Типы жизни, основанные на окислительном фосфорилировании (139К)
IV. Мир прокариот: архебактерии
o Глава 17. Архебактерии: общая характеристика
o Глава 18. Группы архебактерий (49К)
V. Заключение
Литература
В микробиологических учебниках можно найти много интересного. Огромный мир микроскопических живых существ — вот предмет микробиологии. Взгляд любого человека на жизнь как явление, взгляд биолога, изучающего эту жизнь, будет ограничен, если не учитывать жизнь микроскопического мира.
Существуют многие подходы к изучению микроорганизмов, утонченные методики, разнообразные способы анализа, схемы определителей для тех, кто интересуется выделением новых видов, и многое другое. Написаны учебники, предназначенные специально для микробиологов и посвященные различным разделам микробиологии, которая сейчас все больше и больше становится фундаментальной основой многих прикладных дисциплин, таких как биотехнология, медицина и т. д.
Учебник, который предлагается вашему вниманию, как нам кажется, может быть использован и микробиологами, но в первую очередь предназначается для биологов различных профилей, потому что в нем подчеркивается своеобразие способов существования микроорганизмов, которые порой отличаются друг от друга тем, как взаимодействуют с окружающей природой, гораздо больше, чем по внешнему их виду.
Объекты микробиологии объединяют, прежде всего, их чрезвычайно небольшие размеры. Это создает трудности при изучении и диктует необходимость специальных методов наблюдения, но в то же время позволяет микроорганизмам существовать в тонких пленочках вокруг частиц почвы, каплях воды, микроскопических щелях в горных породах, т. е. таких микроэкологических условиях, которые порой, коренным образом, отличаются от макроэкологических условий, характерных для нашей планеты в целом.
Способность микроорганизмов существовать в этих условиях, возможно, позволила им сохранить свойства, соответствующие далекому прошлому нашей Земли. Поэтому мы имеем дело как бы с "миром ископаемых". Для биолога это обстоятельство представляется чрезвычайно важным, потому что если он в своих рассуждениях о жизни не будет учитывать тех особенностей, которые характерны для микроорганизмов, взгляд биолога на жизнь будет слишком узким. Он не будет соответствовать тем возможностям, которыми жизнь на самом деле располагает и которые, может быть, являются отголосками прошедшей эволюции, но сохранились до наших времен именно потому, что сохранились и существуют микроскопические живые объекты.
Итак, этот учебник не о пользе, которую приносят микробы, не о том вреде, который они также могут приносить, что само по себе очень важно и излагается в других учебных пособиях. В этом учебнике мы старались подчеркнуть прежде всего биологическое значение микроорганизмов, подтверждающих своим существованием теорию биохимического единства жизни и одновременно иллюстрирующих возможности далеко идущего разнообразия на уровне физиологии и экологических возможностей. Вот это, как нам кажется, основная идея учебника, полезная для биологов разных профилей. И нам представляется, что третье издание, в котором мы уже можем описать не только некие новые формы, но даже новое царство живого, базирующееся на изучении микроорганизмов, это издание может быть адресовано не только и не столько микробиологам, студентам и аспирантам, но и научным сотрудникам вообще. Мы надеемся, что эта цель нами достигнута, насколько это возможно.
М. В. Гусев, Л. А. Минеева
I. Вводная часть
ГЛАВА 1. ИСТОРИЧЕСКИЙ ОЧЕРК
На протяжении длительного времени человек жил в окружении невидимых существ, использовал продукты их жизнедеятельности (например, при выпечке хлеба из кислого теста, приготовлении вина и уксуса), страдал, когда эти существа являлись причинами болезней или портили запасы пищи, но не подозревал об их присутствии. Не подозревал потому, что не видел, а не видел потому, что размеры этих микросуществ лежали много ниже того предела видимости, на который способен человеческий глаз. Известно, что человек с нормальным зрением на оптимальном расстоянии (25–30 см) может различить в виде точки предмет размером 0,07–0,08 мм. Меньшие объекты человек заметить не может. Это определяется особенностями строения его органа зрения.
Попытки преодолеть созданный природой барьер и расширить возможности человеческого глаза были сделаны давно. Так, при археологических раскопках в Древнем Вавилоне находили двояковыпуклые линзы — самые простые оптические приборы. Линзы были изготовлены из отшлифованного горного хрусталя. Можно считать, что с их изобретением человек сделал первый шаг на пути в микромир.
Дальнейшее совершенствование оптической техники относится к XVI–XVII вв. и связано с развитием астрономии. В это время голландские шлифовальщики стекла сконструировали первые подзорные трубы. Оказалось, что если линзы расположить не так, как в телескопе, то можно получить увеличение очень мелких предметов. Микроскоп подобного типа был создан в 1610 г. Г. Галилеем (G. Galilei, 1564—1642). Изобретение микроскопа открыло новые возможности для изучения живой природы.
Одним из первых микроскоп, состоящий из двух двояковыпуклых линз. дававших увеличение примерно в 30 раз, сконструировал и использовал для изучения строения растений английский физик и изобретатель Р. Гук (R. Hooke, 1635—1703). Рассматривая срезы пробки, он обнаружил правильное ячеистое строение древесной ткани. Эти ячейки впоследствии были названы им "клетками" и изображены в книге "Микрография" (1665). Именно Р. Гук ввел термин "клетка" для обозначения тех структурных единиц, из которых построен сложный живой организм. Дальнейшее проникновение в тайны микромира неразрывно связано с совершенствованием оптических приборов.
ОТКРЫТИЕ МИКРООРГАНИЗМОВ
Первым человеком, увидевшим микроорганизмы, был голландец Антони ван Левенгук (Antony van Leeuwenhoek, 1632—1723), мануфактурщик из Дельфта1. Заинтересовавшись строением льняного волокна, он отшлифовал для себя несколько грубых линз. Позднее А. ван Левенгук увлекся этой тонкой и кропотливой работой и достиг большого совершенства в деле изготовления линз, названных им "микроскопиями". По внешней форме это были одинарные двояковыпуклые стекла, оправленные в серебро или латунь (то, что мы теперь называем "лупы"), однако по своим оптическим свойствам линзы А. ван Левенгука, дававшие увеличение в 200—270 раз, не знали себе равных. (Достаточно напомнить, что теоретический предел увеличения двояковыпуклой линзы — 250–300 раз.)

А. ван Левенгук
1 По мнению В. Л. Омелянского. "первым исследователем. перед изумленным взором которого открылся таинственный и полный чудес мир микроорганизмов. был ученый иезуит Афанасий Kиpxep (A. Kircher. 1601— 1680), автор ряда сочинений астрологического характера. С помощью довольно сильной лупы он наблюдал мельчайших "червячков" в загнившем мясе, молоке, уксусе, сыре и в крови больных, предполагая, что все это живое население произошло из безжизненных органических материалов" (В. Л. Омелянский. Основы микробиологии. М., 1926. С. 16).
Обладая природной любознательностью, А. ван Левенгук с интересом рассматривал все, что попадалось под руку: воду из пруда, зубной налет, настой перца, слюну, кровь и многое другое. Результаты своих наблюдений он начал посылать в Лондонское Королевское общество, членом которого впоследствии был избран. Всего А. ван Левенгук написал в это общество свыше 170 писем, а позднее завещал ему 26 своих знаменитых "микроскопий". Вот выдержка из одного письма: "24 апреля 1676 г. я посмотрел на... воду под микроскопом и с большим удивлением увидел в ней огромное количество мельчайших живых существ. Некоторые из них в длину были раза в 3–4 больше, чем в ширину, хотя они и не были толще волосков, покрывающих тело вши... Другие имели правильную овальную форму. Был там еще и третий тип организмов — наиболее многочисленный — мельчайшие существа с хвостиками". Сопоставив описание, приведенное в этом отрывке, и оптические возможности имевшихся в распоряжении А. ван Левенгука линз, можно сделать заключение, что в 1676 г. ему впервые удалось увидеть бактерии (рис. 1).

Рис. 1. Рисунок бактерий А. ван Левенгука
А. ван Левенгук повсюду обнаруживал микроорганизмы и пришел к выводу, что окружающий мир густо заселен микроскопическими обитателями. Все виденные им микроорганизмы, в том числе и бактерии, А. ван Левенгук считал маленькими животными, названными им "анималькулями", и был убежден, что они устроены так же, как и крупные организмы, т. е. имеют органы пищеварения, ножки, хвостики и т. д. Открытия А. ван Левенгука были настолько неожиданными и даже фантастическими, что на протяжении почти 50 последующих лет вызывали всеобщее изумление. Будучи в Голландии, Петр I посетил А. ван Левенгука и беседовал с ним. Из этой поездки Петр I привез в Россию микроскоп, а позднее в мастерских при его дворе были изготовлены первые отечественные микроскопы. Дальнейшее систематическое изучение окружающей природы с помощью совершенствовавшихся микроскопов подтверждало обнаруженное А. ван Левенгуком повсеместное распространение микроорганизмов. Три основные проблемы, волновавшие умы ученых на протяжении длительного времени, послужили могучим стимулом для развития исследовании, приведших к возникновению и последующему интенсивному развитию микробиологии: природа процессов брожения и гниения, причины возникновения инфекционных болезней и проблема самозарождения организмов 2.
2 О развитии представлений по проблеме самозарождения см. стр 182.
РАЗВИТИЕ ПРЕДСТАВЛЕНИЙ О ПРИРОДЕ ПРОЦЕССОВ БРОЖЕНИЯ И ГНИЕНИЯ
Многие процессы, осуществляемые микроорганизмами, были известны человеку с незапамятных времен. В первую очередь это гниение и брожение. В сочинениях древних греческих и римских авторов можно найти рецепты приготовления вина, кислого молока, хлеба, свидетельствующие о широком использовании в быту брожений. В средние века алхимики не обошли вниманием эти процессы и изучали их наряду с другими чисто химическими превращениями. Именно в этот период были сделаны первые попытки выяснить природу процессов брожения.
Термин "брожение" (fermentatio) для обозначения всех процессов, идущих с выделением газа, впервые употребил голландский алхимик Я. Б. ван Гельмонт (J. В. van Helmont, 1577— 1644). Позднее брожения стали выделять из группы химических процессов, сопровождающихся газовыделением. Для обозначения материальной движущей силы брожения, его активного начала использовали термин "фермент". Взгляд на брожение и гниение как на чисто химические процессы был сформулирован в 1697 г. немецким врачом и химиком Г. Э. Шталем, (G. Е. Stahl, 1660—1734). По представлениям Г. Шталя, брожение и гниение—это химические превращения, идущие под влиянием молекул "фермента", которые передают присущее им внутреннее активное движение молекулам сбраживаемого субстрата, т. е. выступают в качестве своеобразных катализаторов реакции. Однако эта точка зрения принималась не всеми исследователями.
Одна из первых догадок о связи описанных А. ван Левенгуком "глобул" (дрожжей) с явлениями брожения и гниения принадлежит французскому натуралисту Ж. Л. Л. Бюффону (G. L. L. Buffon, 1707—1788). Весьма близко подошел к пониманию роли дрожжей в процессе брожения французский химик А. Л. Лавуазье (A. L. Lavoisier, 1743—1794), изучавший количественно химические превращения сахара при спиртовом брожении. В 1793 г. он писал: "Достаточно немного пивных дрожжей, чтобы... дать первый толчок к брожению: оно потом продолжается само собой. Я доложу в другом месте о действии фермента в целом". Однако сделать это ему не удалось: А. Лавуазье стал жертвой террора французской буржуазной революции.
С 30-х гг. XIX в. начинается период интенсивных микроскопических наблюдений. В 1827 г. французский химик Ж. Б. Демазьер (J. В. Demazier, 1783—1862) описал строение организмов (дрожжей), формирующих пленку на поверхности пива. Однако в работе Ж. Б. Демазьера нет никаких указаний на возможную связь процесса брожения с развивающейся на поверхности бродящей жидкости пленкой. Спустя 10 лет французский ботаник Ш. Каньяр де Латур (Ch. Cagniard de Latour, 1777—1859) предпринял тщательное микроскопическое изучение осадка, образующегося при спиртовом брожении, и пришел к выводу, что он состоит из живых существ, жизнедеятельность которых и является причиной брожения. Почти одновременно немецкий естествоиспытатель Ф. Кютцинг (F. Kuthzing, 1807—1893), исследуя образование уксуса из спирта, обратил внимание на слизистую массу, имеющую вид пленки на поверхности жидкости. Изучая эту массу, Ф. Кютцинг установил, что она состоит из микроскопических живых организмов и имеет непосредственное отношение к накоплению уксуса в среде. К аналогичным выводам пришел другой немецкий естествоиспытатель Т. Шванн (Th. Schwann, 1810—1882). Таким образом, Ш. Каньяр де Латур, Ф. Кютцинг и Т. Шванн независимо друг от друга и почти одновременно пришли к заключению о связи процессов брожения с жизнедеятельностью микроскопических живых существ.
Однако идеи о биологической природе "фермента" брожения, высказанные тремя исследователями, не получили признания. Более того, они были подвергнуты суровой критике со стороны приверженцев теории физико-химической природы брожения, обвинивших своих научных противников в "легкомыслии в выводах" и отсутствии каких-либо доказательств, подтверждающих эту "странную гипотезу". Господствовавшей оставалась теория физико-химической природы процессов брожения.
ФОРМИРОВАНИЕ ПРЕДСТАВЛЕНИЙ О МИКРОБНОЙ ПРИРОДЕ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИИ
Еще древнегреческий врач Гиппократ (ок. 460—377 до н. э.) высказывал предположение о том, что заразные болезни вызываются невидимыми живыми существами. Авиценна (ок. 980— 1037) в "Каноне медицины" писал о "невидимых" возбудителях чумы, оспы и других заболеваний. Подобные мысли можно обнаружить и в трудах итальянского врача, астронома и поэта Дж. Фракастро (J. Fracastro, 1478—1553). В том, что инфекционные болезни вызываются живыми микроскопическими существами, был глубоко убежден русский врач-эпидемиолог Д. С. Самойлович (1744—1805), пытавшийся под микроскопом обнаружить возбудителя чумы, однако возможности существовавших тогда микроскопов не позволили ему этого сделать. В 1827 г. итальянский естествоиспытатель А. Басси (A. Bassi, 1773—1856), изучая заболевание шелковичных червей, обнаружил передачу болезни при переносе микроскопического грибка от больной особи к здоровой. Таким образом, А. Басси впервые удалось экспериментально установить микробную природу этого заболевания.
Несмотря на блестящие догадки отдельных ученых и опыты А. Басси. в целом представление о микробной природе инфекционных болезней в течение долгого времени не получало признания. Подавляющее большинство исследователей были убеждены в том, что причинами всех заболеваний являются нарушения течения химических процессов в организме. Однако острый интерес к изучению инфекционных заболеваний и совершенствование микроскопической техники приводили к быстрому накоплению данных, говорящих об участии микробов в инфекционных заболеваниях.
НАУЧНАЯ ДЕЯТЕЛЬНОСТЬ Л. ПАСТЕРА
Человеком, который своими работами положил начало современной микробиологии, был выдающийся французский ученый Луи Пастер (Louis Pasteur, 1822—1895). Научная деятельность Л. Пастера многогранна и охватывала все основные проблемы того времени, связанные с жизнедеятельностью микроорганизмов.
Чтобы оценить гигантский научный труд Л. Пастера, достаточно привести надпись на доске, прибитой к дому, где помещалась его лаборатория. Надпись эта гласит: "Здесь была лаборатория Л. Пастера:
1857 г. — Брожения. 1860 г. - Самопроизвольное зарождение.
1865 г. — Болезни вина и пива.
1868 г. — Болезни шелковичных червей.
1881 г. — Зараза и вакцина. 1885 г. — Предохранение от бешенства".
Трудно переоценить значение научных открытий Л. Пастера, каждого из которых достаточно, чтобы навсегда вписать имя ученого в историю науки. Изучая молочнокислое, спиртовое, маслянокислое брожение, Л. Пастер выяснил, что эти процессы вызываются определенными видами микроорганизмов и непосредственно связаны с их жизнедеятельностью. Позднее, изучая "болезни" вина, болезни животных и человека, он экспериментально установил, что их "виновниками" также являются микроорганизмы. Таким образом, Л. Пастер впервые показал, что микроорганизмы — это активные формы, полезные или вредные, энергично воздействующие на окружающую природу. в том числе и на человека.
 |
| Л. Пастер |
Принципиально важным не только для микробиологии, но для более глубокого понимания сущности живого в его разнообразных проявлениях было открытие Л. Пастером у микроорганизмов новых типов жизни, не похожих на те, которые имеют место в мире растений и животных. В 1857 г. Л. Пастер при изучении спиртового брожения установил, что оно — результат жизнедеятельности дрожжей без доступа кислорода. Позднее при изучении маслянокислого брожения он обнаружил, что возбудители брожения вообще отрицательно относится к кислороду и могут размножаться только в условиях, исключающих его свободный доступ. Таким образом, Л. Пастер обнаружил существование "жизни без кислорода", т. е. анаэробный способ существования. Он же ввел термины "аэробный" и "анаэробный" для обозначения жизни в присутствии или в отсутствие молекулярного кислорода.
К области теоретических открытий Л. Пастера относятся его работы о невозможности самозарождения. Спор о том, откуда возникают живые существа, в том числе и микроорганизмы: из себе подобных или из других компонентов живой природы, — это давний спор, приобретший к середине XIX в. большую остроту и далеко вышедший за рамки чисто научных дискуссий. На основании проделанных экспериментов Л. Пастер пришел к следующему выводу: "Нет, сегодня не имеется ни одного известного факта, с помощью которого можно было бы утверждать, что микроскопические существа появились на свет без зародышей, без родителей, которые их напоминают. Те, кто настаивает на противоположном, являются жертвой заблуждения или плохо проделанных опытов, содержащих ошибки, которые они не сумели заметить или которых они не сумели избегнуть".
И наконец, работы Л. Пастера в области изучения инфекционных болезней животных и человека (болезнь шелковичных червей, сибирская язва, куриная холера, бешенство) позволили ему не только выяснить природу этих заболеваний, но и найти способ борьбы с ними. Поэтому мы с полным правом можем считать, что своими классическими работами по изучению инфекционных болезней и мер борьбы с ними Л. Пастер положил начало развитию медицинской микробиологии.
Работы Л. Пастера были по достоинству оценены его современниками и получили международное признание. В 1888 г. для ученого на средства, собранные по международной подписке, был построен в Париже научно-исследовательский институт, носящий в настоящее время его имя. Л. Пастер был первым директором этого института. Открытия Л. Пастера показали, как разнообразен, необычен, активен невидимый простым глазом микромир и какое огромное поле деятельности представляет его изучение.
УСПЕХИ МИКРОБИОЛОГИИ ВО ВТОРОЙ ПОЛОВИНЕ XIX В.
Оценивая успехи, достигнутые микробиологией во второй половине XIX в., французский исследователь П. Таннери (Р. Tannery) в работе "Исторический очерк развития естествознания в Европе (1300—1900)" писал: "Перед лицом бактериологических открытий история других естественных наук за последние десятилетия XIX столетия кажется несколько бледной". Успехи микробиологии в этот период непосредственно связаны с новыми идеями и методическими подходами, внесенными в микробиологические исследования Л. Пастером. В числе первых, кто оценил значение открытий Л. Пастера, был английский хирург Дж. Листер (J. Lister, 1827—1912). Он понял, что причина большого процента смертных случаев после операций — заражение ран бактериями из-за незнания, во-первых, и несоблюдения, во-вторых, элементарных правил антисептики. Дж. Листер впервые ввел в медицинскую практику методы предупреждения подобного заражения ран, заключавшиеся в обработке всех хирургических инструментов карболовой кислотой и разбрызгивании ее в операционной во время операции. Таким путем он добился существенного снижения числа смертельных исходов после операций.
Одним из основоположников медицинской микробиологии наряду с Л. Пастером явился немецкий микробиолог Р. Кох (R. Koch, 1843—1910), занимавшийся изучением возбудителей инфекционных заболеваний. Свои исследования Р. Кох начал, еще будучи сельским врачом, с изучения сибирской язвы и в 1877 г. опубликовал работу, посвященную возбудителю этого заболевания — Bacillus anthracis. Вслед за этим внимание Р. Коха привлекла другая тяжелая и широко распространенная болезнь того времени—туберкулез. В 1882 г. Р. Кох сообщил об открытии возбудителя туберкулеза, который в его честь был назван "палочкой Коха". (В 1905 г. за исследование туберкулеза Р. Коху была присуждена Нобелевская премия.) Ему принадлежит также открытие возбудителя холеры.
Родоначальником русской микробиологии является Л. С. Ценковский (1822—1887). Объектом его исследований были микроскопические простейшие, водоросли, грибы. Л. С. Ценковский открыл и описал большое число простейших, изучал их морфологию и циклы развития. Это позволило ему сделать вывод об отсутствии резкой границы между миром растений и животных. Л. С. Ценковский интересовался проблемами медицинской микробиологии. Им была организована одна из первых Пастеровских станций в России и предложена вакцина против сибирской язвы (так называемая "живая вакцина Ценковского").
Основоположником медицинской микробиологии справедливо считают также И. И. Мечникова (1845—1916). И. И. Мечников был разносторонним исследователем, но основные свои научные интересы он сосредоточил на проблеме изучения взаимоотношений хозяина и микроорганизма-паразита. В 1883 г. И. И. Мечников создал фагоцитарную теорию иммунитета. Невосприимчивость человека к повторному заражению после перенесенного инфекционного заболевания была известна давно. Однако природа этого явления оставалась непонятной и после того, как были разработаны и широко применялись прививки против ряда инфекционных заболеваний. И. И. Мечников показал, что защита организма от болезнетворных микроорганизмов — сложная биологическая реакция, в основе которой лежит способность белых кровяных телец (фагоцитов) захватывать и разрушать посторонние тела, попавшие в организм. Вклад И. И. Мечникова в науку был оценен его современниками. В 1909 г. за исследования по фагоцитозу И. И. Мечникову была присуждена Нобелевская премия.
 |
| С. Н. Виноградский |
Большой вклад в развитие общей микробиологии внесли русский микробиолог С. Н. Виноградский (1856—1953) и голландский микробиолог М. Бейеринк (М. Beijerinck, 1851—1931). Оба много и плодотворно работали в разных областях микробиологии. Впитав идеи Л. Пастера о многообразии форм жизни в микромире, С. Н. Виноградский ввел микроэкологический принцип в исследование микроорганизмов.
Для выделения в лабораторных условиях группы бактерий с определенными свойствами С. Н. Виноградский предложил создавать специфические (элективные) условия, дающие возможность преимущественного развития данной группы организмов. Поясним это примером. С. Н. Виноградский предположил, что среди микроорганизмов есть виды, способные усваивать молекулярный азот атмосферы, являющийся инертной формой азота по отношению ко всем животным и растениям. Для выделения таких микроорганизмов в питательную среду были внесены источники углерода, фосфора и другие минеральные соли, но не добавлено никаких соединений, содержащих азот. В результате в этих условиях не могли расти микроорганизмы, которым необходим азот в форме органических или неорганических соединений, но могли расти виды, обладавшие способностью фиксировать азот атмосферы. Именно так С. Н. Виноградским в 1893 г. был выделен из почвы анаэробный азотфиксатор, названный им в честь Л. Пастера Clostridium pasteurianum.
Пользуясь изящными методическими приемами, в основу которых был положен микроэкологический принцип, С. Н. Виноградский выделил из почвы микроорганизмы, представляющие собой совершенно новый тип жизни и получившие название хемолитоавтотрофных. В качестве единственного источника углерода для построения всех веществ клетки хемолитоавтотрофы используют углекислоту, а энергию получают в результате окисления неорганических соединений серы, азота, железа, сурьмы или молекулярного водорода.
 |
| М. Бейеринк |
Микроэкологический принцип был успешно развит М. Бейеринком и применен при выделении различных групп микроорганизмов. В частности, спустя восемь лет после открытия С. Н. Виноградским анаэробного азотфиксатора, М. Бейеринк обнаружил в почве еще один вид бактерий, способных к росту и азотфиксации в аэробных условиях, — Azotobacter chroococcum. Круг научных интересов М. Бейеринка был необычайно широк. Ему принадлежат работы по исследованию физиологии клубеньковых бактерий, изучению процесса денитрификации и сульфатредукции, работы по изучению ферментов разных групп микроорганизмов.
С. Н. Виноградский и М. Бейеринк являются основоположниками экологического направления микробиологии, связанного с изучением роли микроорганизмов в природных условиях и участием их в круговороте веществ в природе.
Сообщения об активном участии микроорганизмов в процессах превращения веществ в природе стали быстро накапливаться в 70—80-х гг. XIX в. В 1877 г. французские химики Т. Шлезинг (Т. Schloesing) и А. Мюнц (A. Muntz) доказали микробиологическую природу процесса нитрификации. В 1882 г. П. Дегерен (Р. Deherein) обнаружил аналогичную природу процесса денитрификации, а двумя годами позднее он же установил микробиологическую природу анаэробного разложения растительных остатков. М. С. Воронин в 1867 г. описал клубеньковые бактерии, а спустя почти двадцать лет Г. Гельригель (Н. Неllrigеl) и Г. Вильфарт (Н. Willfarth) показали их способность к азотфиксации. П. А. Костычев создал теорию микробиологической природы процессов почвообразования. Конец XIX в. ознаменовался еще одним важным открытием в области микробиологии. В 1892 г. Д. И. Ивановский обнаружил вирус табачной мозаики — представителя новой группы микроскопических существ. В 1898 г. независимо от Д. И. Ивановского вирус табачной мозаики был описан М. Бейеринком.
Таким образом, вторая половина XIX в. характеризуется выдающимися открытиями в области микробиологии. На смену описательному морфолого-систематическому изучению микроорганизмов, господствовавшему в первой половине XIX в., пришло физиологическое изучение микроорганизмов, основанное на точном эксперименте. Развитие нового этапа микробиологии связано в первую очередь с трудами Л. Пастера. К концу XIX в. намечается дифференциация микробиологии на ряд направлений: общая, медицинская, почвенная.
МИКРОБИОЛОГИЯ В XX В.
Успехи микробиологии во второй половине XIX в. привели к обнаружению чрезвычайного разнообразия типов жизни в микромире. Следующий вопрос, заинтересовавший исследователей: как объяснить такое многообразие, определить его границы, выявить, на чем оно основано? Постановкой этой проблемы, имеющей общебиологическое значение, мы обязаны двум крупнейшим микробиологам нашего времени А. Клюйверу (А. Kluyver 1888—1956) и К. ван Нилю (С. van Niel, 1897–1985). А. Клюйвер и его ученики (одним из них был К. ван Ниль) провели сравнительные биохимические исследования в относительно далеко отстоящих друг от друга физиологических группах микроорганизмов. Было изучено много форм микроорганизмов и примерно к середине 50-х гг. нашего века сформулировано то, что теперь называют теорией биохимического единства жизни.
В чем же конкретно состоит биохимическое единство жизни? Общее основано на единстве конструктивных, энергетических процессов и механизмов передачи генетической информации. А. Клюйвер доказал два первых положения: все живые организмы построены из однотипных химических макромолекул, универсальной единицей биологической энергии служит АТФ, в основе физиологического разнообразия живых существ лежит несколько основных метаболических путей. Что касается последнего положения, то А. Клюйвер изучением этой проблемы не занимался. Единство системы передачи генетической информации у всех клеточных типов жизни было установлено позднее. В настоящее время мы пока не знаем исключений, которые ставили бы под сомнение теорию биохимического единства жизни.
С начала XX в. продолжается дальнейшая дифференциация микробиологии. От нее отпочковываются новые научные дисциплины (вирусология, микология) со своими объектами исследования, выделяются направления, различающиеся задачами исследования (общая микробиология, техническая, сельскохозяйственная, медицинская, генетика микроорганизмов). Перечисление достижений микробиологии XX в. в кратком очерке представляется необычайно сложным, что и привело нас к заключению не делать этого. Фактически все последующее изложение материала (и то достаточно краткое и не затрагивающее всех направлений современной микробиологии) есть попытка охарактеризовать достижения в некоторых областях микробиологии на современном этапе. Вклад отдельных исследователей в решение определенных микробиологических проблем мы старались отмечать по мере изложения материала.
Итак, мы коротко остановились на истории микробиологии, особо подчеркнув роль исследователей, работы которых имели этапное значение не только для развития микробиологии, но и биологии в целом: А. ван Левенгук — открытие микромира, Л. Пастер — выяснение роли микроорганизмов в природе, С. Н. Виноградский и М. Бейеринк — утверждение многообразия форм жизни в микромире, А. Клюйвер и К. ван Ниль — доказательство биохимического единства жизни.
ГЛАВА 2. ПОЛОЖЕНИЕ МИКРООРГАНИЗМОВ В СИСТЕМЕ ЖИВОГО МИРА
Начиная с Аристотеля (384—322 до н. э.), которому принадлежит первая попытка систематизировать накопленные к тому времени сведения об организмах, биологи делили живой мир на два царства — растений и животных. А. ван Левенгук, открывший мир микроскопических живых существ, был убежден в том, что они являются "маленькими живыми зверушками". С этого времени и до XIX в. все открываемые микроорганизмы рассматривали как мельчайшие существа животной природы.
Во второй половине XIX в. немецкий биолог Э. Геккель (Е. Haeckel, 1834—1919) приходит к заключению, что микроорганизмы настолько существенно отличаются как от царства животных, так и от царства растений, что не укладываются ни в одно из этих подразделений. Э. Геккель предложил выделить все микроорганизмы, у которых отсутствует дифференцировка на органы и ткани (простейшие, водоросли, грибы, бактерии), в отдельное царство Protista (протисты, первосущества)3, включив в него организмы, во многих отношениях занимающие промежуточное положение между растениями и животными. Термин "protista" и сейчас применим для обозначения объектов, исследуемых микробиологами.
3 По-гречески protos — самый простой.
В настоящее время нет единства во взглядах на общую систему живого мира. Согласно одной из точек зрения попытки уложить все существующее разнообразие организмов в жесткую схему нецелесообразны, поскольку любые искусственные разграничения нарушают естественные связи между организмами. Следствие этого — тенденция наименьшего дробления органического мира, признание целесообразности выделения только двух царств: Plantae (растения) и Animalia (животные). Эта точка зрения акцентирует внимание на чертах сходства, соединяющих различные типы организмов, и на существовании переходов от одной группы организмов к другой в процессе эволюции. В соответствии с противоположным представлением разделение всех живых форм на крупные таксоны (царства) наиболее полно отражает существующее многообразие типов жизни, подчеркивая эту сторону живого мира. Согласно первой точке зрения все микроорганизмы рассматриваются как примитивные растения или животные и соответственно входят в состав царств Plantae или Animalia. Согласно второй — микроорганизмы могут претендовать на уникальное место в иерархии живых форм, что впервые понял Э. Геккель. Дальнейшее изучение геккелевских "первосуществ" выявило неоднородность этой группы. Тогда же стало ясно, что понятие "микроорганизм" не имеет таксономического смысла. Оно объединяет организмы по признаку их малых (как правило, видимых только с помощью соответствующих приборов) размеров и связанных с этим специфических методов изучения.
Данные о различии в строении клеток микроорганизмов, входящих в группу Protista, начали накапливаться с конца XIX в, Это повлекло за собой деление группы на высшие и низшие протисты. К высшим протистам стали относить микроскопических животных (простейших), микроскопические водоросли (кроме сине-зеленых) и микроскопические грибы (плесени, дрожжи), к низшим — все бактерии и сине-зеленые водоросли (последние чаще называют теперь цианобактериями). Деление на высшие и низшие протисты происходило в соответствии с двумя выявленными типами клеточной организации — эукариотной и прокариотной4. Высшие протисты имеют эукариотное строение клеток, т. е. являются эукариотами, низшие — прокариотное.
4 Термины были предложены в 30-х гг. XX в. протозоологом Э. Шаттоном (Е. Chatton).
Обоснование того, что прокариотный и эукариотный типы клеточной организации являются наиболее существенной границей, разделяющей все клеточные формы жизни, связано с работами Р. Стейниера (R. Stanier, 1916—1982) и К. ван Ниля, относящимися к 60-м гг. Поясним разницу между прокариотами и эукариотами. Клетка — это кусочек цитоплазмы, отграниченный мембраной. Последняя под электронным микроскопом имеет характерную ультраструктуру: два электронно-плотных слоя каждый толщиной 2,5—3,0 нм, разделенных электронно-прозрачным промежутком. Такие мембраны получили название элементарных. Обязательными химическими компонентами каждой клетки являются два вида нуклеиновых кислот (ДНК и РНК), белки, липиды, углеводы. Цитоплазма и элементарная мембрана, окружающая ее, — непременные и обязательные структурные элементы клетки. Это то, что лежит в основе строения всех без исключения клеток. Изучение тонкой структуры выявило существенные различия в строении клеток прокариот (бактерий и цианобактерий) и эукариот (остальные макро- и микроорганизмы).
Прокариотная клетка отличается тем, что имеет одну внутреннюю полость, образуемую элементарной мембраной, называемой клеточной, или цитоплазматической (ЦПМ). У подавляющего большинства прокариот ЦПМ — единственная мембрана, обнаруживаемая в клетке. В эукариотных клетках в отличие от прокариотных есть вторичные полости. Ядерная мембрана, отграничивающая ДНК от остальной цитоплазмы, формирует вторичную полость. Наружные мембраны хлоропластов и митохондрий, окружающие заключенные в них функционально специализированные мембраны, играют аналогичную роль. Клеточные структуры, ограниченные элементарными мембранами и выполняющие в клетке определенные функции, получили название органелл. Ядро, митохондрий, хлоропласты — это клеточные органеллы. В эукариотных клетках помимо перечисленных выше есть и другие органеллы.
В клетках прокариот органеллы, типичные для эукариот, отсутствуют. Ядерная ДНК у них не отделена от цитоплазмы мембраной. В цитоплазме находятся функционально специализированные структуры, но они не изолированы от цитоплазмы с помощью мембран и, следовательно, не образуют замкнутых полостей. Эти структуры могут быть сформированы и мембранами, но последние не замкнуты и, как правило, обнаруживают тесную связь с ЦПМ, являясь результатом ее локального внутриклеточного разрастания. В клетках прокариот есть также образования, окруженные особой мембраной, имеющей иное по сравнению с элементарной строение и химический состав.
Таким образом, основное различие между двумя типами клеток — существование в эукариотной клетке вторичных полостей, сформированных с участием элементарных мембран. Сопоставление некоторых черт клеточной организации прокариотных и эукариотных организмов представлено в табл. 1.
Таблица 1. Сопоставление некоторых черт прокариотной и эукариотной клеточной организации
| Признак | Прокариотная клетка | Эукариотная клетка |
| Организация генетического материала | нуклеоид (ДНК не отделена от цитоплазмы мембраной), состоящий из одной хромосомы; митоз отсутствует | ядро (ДНК отделена от цитоплазмы ядерной оболочкой), содержащее больше одной хромосомы; деление ядра путем митоза |
| Локализация ДНК | в нуклеоиде и плазмидах, не ограниченных элементарной мембраной | в ядре и некоторых органеллах |
| Цитоплазматические органеллы | отсутствуют | имеются |
| Рибосомы в цитоплазме | 70S-типа | 80S-типа |
| Цитоплазматические органеллы | отсутствуют | имеются |
| Движение цитоплазмы | отсутствует | часто обнаруживается |
| Клеточная стенка (там, где она имеется) | в большинстве случаев содержит пептидогликан | пептидогликан отсутствует |
| Жгутики | нить жгутика построена из белковых субъединиц, образующих спираль | каждый жгутик содержит набор микротрубочек, собранны в группы: 2·9-2 |
В связи с тем что прокариотная и эукариотная организация клеток принципиально различна, было предложено только на основании этого признака выделить все прокариоты в особое царство. Р. Меррей (R. Murray) в 1968 г. предложил все клеточные организмы разделить на две группы по ТИПУ их клеточной организации: царство Prokaryotae, куда вошли все организмы с прокариотным строением клетки, н царство Eukaryotae, куда включены все высшие протисты, растения и животные.
Р. Виттэкер (R. Whittaker) предложил схему, по которой все живые организмы, имеющие клеточное строение, представлены разделенными на пять царств (рис. 2). Такая система классификации живого мира отражает три основных уровня его клеточной организации: Monera включает прокариотные организмы, находящиеся на самом примитивном уровне клеточной организации; Protista — микроскопические, в большинстве своем одноклеточные, недифференцированные формы жизни, сформировавшиеся в результате качественного скачка в процессе эволюции, приведшего к возникновению эукариотных клеток; многоклеточные эукариоты представлены в свою очередь тремя царствами Plantae, Fungi и Animalia.
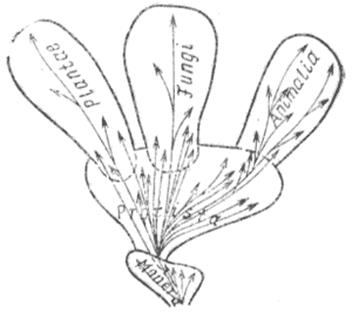
Рис. 2. Схема пяти царств живого мира: прокариоты (царство Monera), одноклеточные эукариоты (царство Protista), многоклеточные эукариоты (царства Plantae, Fungi, Animalia) (no Whittaker, 1969)
Три последние таксономические группы различаются по способу питания: фототрофный тип питания за счет процесса фотосинтеза характерен для растений (Plantae): грибы (Fungi) в основном характеризуются осмотрофным типом питания, т. е. питанием растворенными органическими веществами; животные (Animalia) осуществляют голозойное питание, заключающееся в захватывании и переваривании твердой пищи. Способы питания, специфические для растений и грибов, возникли в процессе эволюции на уровне Monera. На уровне Protista они получили свое дальнейшее развитие; здесь же сформировался третий тип питания — голозойный.
Не берясь судить о целесообразности деления живой природы на пять или шесть царств, можно с определенностью утверждать, что обособление прокариотных микроорганизмов в отдельное царство Prokaryotae правомерно, поскольку основано на принципиальных различиях в структуре прокариотных и эукариотных клеток, т. е. тех единиц, из которых построены все клеточные формы жизни.
ГЛАВА 3. РАЗМЕРЫ МИКРООРГАНИЗМОВ
Как показывает само название, объекты, относимые к микроорганизмам, были выделены по признаку их малых размеров. Если принять за критерий границу видимости невооруженным глазом, равную 70—80 мкм5, то все объекты, которые лежат за пределами этой границы, можно отнести к микроорганизмам. Мир микроорганизмов — это преимущественно мир одноклеточных форм. Диапазон размеров микроорганизмов велик (табл.2). Величина самых крупных представителей микромира, лежащих на границе видимости невооруженным глазом, приблизительно 100 мкм (некоторые диатомовые водоросли, высшие протисты). На порядок ниже размеры одноклеточных зеленых водорослей и клеток дрожжей, еще ниже размеры, характерные для большинства бактерий. В среднем линейные размеры бактерий лежат в пределах 0,5–3 мкм, но есть среди бактерий свои "гиганты" и "карлики". Например, клетки нитчатой серобактерии Beggiatoa alba имеют диаметр до 50 мкм; Achromatium oxaliferum, считающийся одним из крупных бактериальных организмов, имеет в длину 15—100 мкм при поперечнике примерно 5—33 мкм, а длина клетки спирохеты может быть до 250 мкм.
5 1 миллиметр (мм) = 103 микрометров (мкм) = 106 нанометров (нм) = 107 ангстрем (Å) = 1011 пикометров (пм).
Таблица 2. Размеры различных объектов
| Объект | Линейные размеры, мкм* |
| Одноклеточные эукариоты | |
| Некоторые диатомовые водоросли и высшие протисты | |
| Зеленая водоросль Chlorella | 2-10 |
| Клетка дрожжей Saccharomyces | 6-10 |
| Прокариотные организмы | |
| Крупные | |
| Achromatium oxaliferum | 5-33х15-100 |
| Beggiatoa alba | 2-10х1-50 |
| Cristispira pectinis | 1,5х36-72 |
| Macromonas mobilis | 6-14х10-30 |
| Thiovulum majus | 5-25 |
| Spirochaeta plicatilis | 0,2-0,7х80-250 |
| Обычные | |
| Bacillus subtilis | 0,7-0,8x2-3 |
| Escherichia coli | 0,3-1х1-6 |
| Staphylococcus aureus | 0,5-1,0 |
| Thiobacillus thioparus | 0,5х1-3 |
| Rickettsia prowazeki | 0,3-0,6x0,8-2 |
| Мелкие | |
| Mycoplasma mycoides | 0,1х0,25 |
| Bdellovibrio bacteriororus | 0,3x1,2 |
| Haemobarfonella muris | 0,1x0,3-0,7 |
| Wolbachia melophagi | 0,3х0,6 |
| Вирусы | |
| Крупные | |
| табачной мозаики | 0,02x0,3 |
| коровьей оспы | 0,26 |
| гриппа | 0,1 |
| фаг Т2 | 0,06x0,2 |
| Мелкие | |
| 0Х174 | 0,025 |
| желтой лихорадки | 0,022 |
| вирус-сателлит | 0,018 |
| Толщина ЦПМ бактериальной клетки | 0,01 |
| Рибосома | 0,018 |
| Молекула глобулярного белка | |
| крупная | 0,013 |
| мелкая | 0,004 |
Самые мелкие из известных прокариотных клеток — бактерии, принадлежащие к группе микоплазм. Описаны микоплазмы с диаметром клеток 0,1–0,15 мкм. Поскольку молекулы всех соединений имеют определенные физические размеры, то, исходя из объема клетки с диаметром 0, 15 мкм, легко подсчитать. что в ней может содержаться порядка 1200 молекул белка и осуществляться около 100 ферментативных реакций. Минимальное число ферментов, нуклеиновых кислот и других макромолекулярных компонентов, необходимых для самовоспроизведения теоретической "минимальной клетки", составляет, по про веденной оценке, около 50. Это то, что необходимо для поддержания клеточной структуры и обеспечения клеточного метаболизма. Таким образом, в группе микоплазм достигнут размер клеток, близкий к теоретическому пределу клеточного уровня организации жизни. Мельчайшие микоплазменные клетки равны или даже меньше частиц другой группы микроскопических организмов — вирусов.
Если бактериальные клетки обычно можно увидеть в световой микроскоп, то вирусы, размеры большинства которых находятся в диапазоне 16–200 нм, лежат за пределами его разрешающей способности. Впервые наблюдать вирусы и выяснить их структуру удалось после изобретения электронного микроскопа. По своим размерам вирусы занимают место между самыми мелкими бактериальными клетками и самыми крупными органическими молекулами. Размер частиц вируса-сателлита (18 нм) и величина крупной молекулы глобулярного белка (13 нм) близки. Таким образом, если раньше между известными биологам организмами и неживыми молекулами химиков существовала пропасть, то теперь этой пропасти нет: она заполнена вирусами.
Размеры всех живых организмов, выраженные в одних единицах, например в ангстремах, располагаются в диапазоне от 102 (самые мелкие вирусы) до 1011 (размеры кита). Если за границу, разделяющую микро- и макромиры, принять предел видимости невооруженным глазом, т. е. приблизительно 106Å. то, как можно видеть из приведенных значений, на долю микромира приходится огромный диапазон величин.
Краткое рассмотрение различных представителей микромира, занимающих определенные "этажи" размеров, показывает. что, как правило, величина объектов определенно связана с их структурной сложностью. Нижний предел размеров свободноживушего одноклеточного организма определяется пространством, требуемым для упаковки внутри клетки аппарата, необходимого для независимого существования. Ограничение верхнего предела размеров микроорганизмов определяется, по современным представлениям, соотношениями между клеточной поверхностью и объемом. При увеличении клеточных размеров поверхность возрастает в квадрате, а объем — в кубе, поэтому соотношение между этими величинами сдвигается в сторону последнего. У микроорганизмов по сравнению с макроорганизмами очень велико отношение поверхности к объему. Это создает благоприятные условия для активного обмена между микроорганизмами и внешней средой. И действительно, метаболическая активность микроорганизмов, измеренная по разным показателям, в расчете на единицу биомассы намного выше, чем у более крупных клеток. Поэтому представляется закономерным, что низшие формы жизни могли возникнуть и в настоящее время могут существовать только на базе малых размеров, так как последние создают целый ряд преимуществ, обеспечивающих жизнеспособность этим формам жизни.
ГЛАВА 4. СТРОЕНИЕ ПРОКАРИОТНОЙ КЛЕТКИ
Приведенные в главе 2 схемы деления клеточных организмов на высшем уровне предусматривают выделение всех прокариот в отдельное царство. В 70-х гг. обнаружены микроорганизмы, структурно относящиеся к прокариотному типу, но значительно отличающиеся химическим строением важных клеточных макромолекул и способностью осуществлять уникальные биохимические процессы. Эти необычные прокариотные организмы были названы архебактериями. Типичные прокариоты, или бактерии, получили соответственно название эубактерий (истинных бактерий). Число известных архебактерий по сравнению с эубактериями чрезвычайно мало.
Материал настоящего раздела посвящен общей характеристике прокариотных организмов (в основном эубактерий), отличающихся морфологическим и особенно физиологическим разнообразием. В основе морфологического разнообразия лежат различия в размерах и форме отдельных клеток, способах их деления, природе и наборе цитоплазматических включений, строении клеточной стенки и структур, локализованных снаружи от нее, наличии и типе дифференцированных форм, образующихся в процессе жизненного цикла. Всем этим вопросам посвящены главы 4 и 5. В главах 6—9 представлена общая картина физиологического разнообразия прокариот, складывающегося из различий в механизмах получения энергии и источниках питания, разного отношения к молекулярному кислороду и другим факторам внешней среды, прежде всего свету, температуре, кислотности среды. В главе 10 обсуждаются генетические механизмы, приведшие в процессе эволюции к структурно-физиологическому разнообразию прокариот. Глава 11, посвященная проблемам систематики и описанию основных групп прокариот, иллюстрирует на конкретных примерах материал, представленный в предыдущих главах. Завершает раздел глава 12, в которой излагается наиболее общепринятая гипотеза происхождения жизни на Земле, приведшая к возникновению первичной клетки, и имеющийся в настоящее время экспериментальный материал, подтверждающий эту гипотезу.
ФОРМА ПРОКАРИОТ
До недавнего времени большинство исследователей традиционно считали, что клетки прокариот достаточно однообразны и в подавляющем большинстве имеют форму сферы, цилиндра или спирали. Они бывают одиночными, в иных случаях образуют нити или колонии. Прокариоты сферической формы, называемые кокками, могут после деления не расходиться. Если деление происходит в одной плоскости, образуются пары клеток (диплококки) или цепочки (стрептококки). В том случае, когда деление происходит относительно равномерно в трех взаимно перпендикулярных направлениях и клетки после деления остаются соединенными друг с другом, возникают пакеты правильной формы (сарцины) или колонии сферической формы.
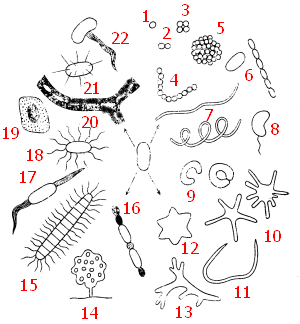
Рис. 3. Разнообразие форм прокариот: 1 — кокк; 2 — диплококк; 3 — сарцина; 4 — стрептококк; 5 — колония сферической формы: 6 — палочковидные бактерии (одиночная клетка и цепочка клеток); 7 — спириллы; 8 — вибрион; 9 — бактерии, имеющие форму замкнутого или незамкнутого кольца; 10 — бактерии, образующие выросты (простеки); 11 — бактерия червеобразной формы; 12 — бактериальная клетка в форме шестиугольной звезды; 13 — представитель актиномицетов; 14 — плодовое тело миксобактерии; 15 — нитчатая бактерия рода Caryophanon с латерально расположенными жгутиками: 16 — нитчатая цианобактерия. образующая споры (акинеты) и гетероцисты; 8, 15, 17, 18 — бактерии с разными типами жгутикования; 19 — бактерии, образующая капсулу; 20 — нитчатые бактерии группы Sphaeroillus, заключенные в чехол, инкрустированный гидратом окиси железа; 21 — бактерия, образующая шипы; 22 — Galionella
Если же деление происходит в нескольких плоскостях неравномерно, образуются клеточные скопления неправильной формы (рис. 3, 1—5). Прокариоты, имеющие форму цилиндра (палочковидные), сильно различаются по величине отношения длины клетки к ее поперечнику. Прокариоты спиралевидной формы характеризуются разным числом витков: у спирилл — от одного до нескольких витков, вибрионы выглядят наподобие изогнутых палочек, так что их можно рассматривать как неполный виток спирали (рис. 3, 6—8).
За последнее время среди прокариот обнаружены организмы, отличающиеся от описанных выше основных форм. Некоторые бактерии имеют вид кольца, замкнутого или разомкнутого в зависимости от стадии роста (рис.3, 9). У прокариот, в основном размножающихся почкованием, описано образование клеточных выростов (простек), число которых может колебаться от 1 до 8 и более (рис. 3, 10). Из природных субстратов выделены бактерии червеобразной формы и напоминающие шестиугольную звезду (рис. 3, 11, 12). Для некоторых видов характерно слабое или довольно хорошо выраженное ветвление (рис. 3, 13). Описаны прокариоты, обладающие морфологической изменчивостью, в зависимости от условий имеющие вид палочек, кокков или обнаруживающие слабое ветвление.
Форма многоклеточных прокариот также разнообразна: это скопления различной конфигурации, чаще — нити (рис. 3, 14—16). Своеобразие бактериальным клеткам придают жгутики, имеющие различное расположение на клеточной поверхности (рис. 3, 8, 15, 17, 18), а также выделения внеклеточных веществ разной химической природы (рис. 3, 19—22).
СТРУКТУРА, ХИМИЧЕСКИЙ СОСТАВ И ФУНКЦИИ КОМПОНЕНТОВ ПРОКАРИОТНОЙ КЛЕТКИ
Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации (рис. 4). Структуры, расположенные снаружи от ЦПМ (клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки), называют обычно поверхностными структурами. Термином "клеточная оболочка" часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). ЦПМ вместе с цитоплазмой называется протопластом. Рассмотрим сначала строение, химический состав и функции поверхностных клеточных структур.
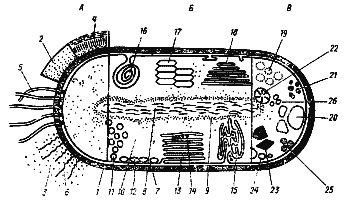
Рис. 4. Комбинированное изображение прокариотной клетки. А — поверхностные клеточные структуры и внеклеточные образования: 1 — клеточная стенка; 2 — капсула; 3 — слизистые выделения; 4 — чехол; 5 — жгутики; 6 — ворсинки; Б — цитоплазматические клеточные структуры: 7 — ЦПМ; 8 — нуклеоид; 9 — рибосомы; 10 — цитоплазма; 11 — хроматофоры; 12 — хлоросомы; 13 — пластинчатые тилакоиды; 14 — фикобилисомы; 15 — трубчатые тилакоиды; 16 — мезосома; 17 — аэросомы (газовые вакуоли); 18 — ламеллярные структуры; В — запасные вещества: 19 — полисахаридные гранулы; 20 — гранулы поли--оксимасляной кислоты; 21 — гранулы полифосфата; 22 — цианофициновые гранулы; 23 — карбоксисомы (полиэдральные тела); 24 — включения серы; 25 — жировые капли; 26 — углеводородные гранулы (по Schlegel, 1972)
Клеточная стенка
Клеточная стенка — важный и обязательный структурный элемент подавляющего большинства прокариотных клеток, располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с окружающей средой (у клеток, не содержащих этих слоев клеточной оболочки). На долю клеточной стенки приходится от 5 до 50% сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. Концентрация солей в клетке, как правило, намного выше, чем в окружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка чисто механически защищает клетку от проникновения в нее избытка воды.
По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты, относящиеся к эубактериям, делятся на две большие группы. Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна: у так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются6. У некоторых эубактерий положительная реакция при окрашивании описанным выше способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки.
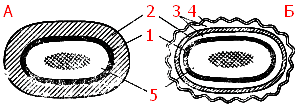
Рис. 5. Клеточная стенка грамположительных (А) и грамотрицательных (Б) эубактерий: 1 — цитоплазматическая мембрана; 2 — пептидогликан; 3 — периплазматическое пространство; 4 — наружная мембрана: 5 — цитоплазма, в центре которой расположена ДНК
Клеточные стенки грамположительных и грамотрицательных эубактерий резко различаются как по химическому составу (табл. 3), так и по ультраструктуре (рис. 5).
В состав клеточной стенки эубактерий входят семь различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке. У грамположительных1 эубактерий он составляет основную массу вещества клеточной стенки (от 40 до 90%), у грамотрицательных — содержание пептидогликана значительно меньше (1—10%). Клеточная стенка цианобактерий, сходная с таковой грамотрицательных эубактерий, содержит от 20 до 50% этого гетерополимера.
1 Этот способ был впервые предложен в 1884 г. датским ученым X. Грамом (Ch. Gram), занимавшимся окрашиванием тканей. Позднее он был использован для бактерий.
Под электронным микроскопом клеточная стенка грамположительных эубактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм. У грамотрицательных эубактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2—3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8—10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных эубактерий получил название наружной мембраны.
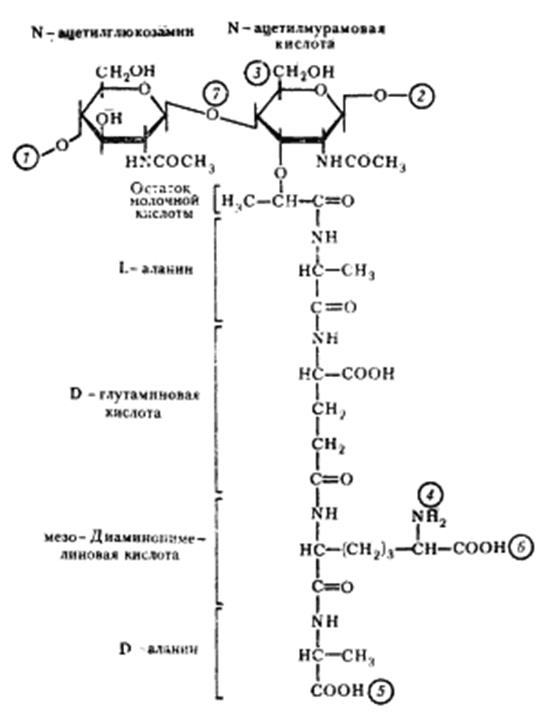
Рис. 6. Структура повторяющейся единицы пептидогликана клеточной стенки эубактерий. Цифры в кружках обозначают: 1, 2— места полимеризации гликанового остова молекулы: 3 — место присоединения с помощью фосфодиэфирной связи молекулы тейхоевой кислоты в клеточной стенке грамположительных эубактерий; 4, 5 — места, по которым происходит связывание между гликановыми цепями с помощью пептидных связей; 6 — место ковалентного связывания (пептидная связь) с липопротеином наружной мембраны у грамотрицательных эубактерий; 7 — место действия лизоцима
Клеточная стенка грамположительных эубактерий плотно прилегает к ЦПМ, в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электронно-прозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической: и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп эубактерий, характерно только для грамотрицательных форм.
Клеточная стенка грамположительных эубактерий. Основную массу клеточной стенки грамположительных эубактерий составляет специфический гетерополимер — пептидогликан. Полисахаридный остов молекулы построен из чередующихся остатков N -ацетилглюкозамина и N -ацетилмурамовой кислоты, соединенных между собой посредством) -1,4-гликозидных связей (рис. 6). К N -ацетилмурамовой кислоте присоединен короткий пептидный хвост, состоящий из небольшого числа (обычно 4—5) аминокислот. У грамположительных эубактерий обнаружено более 100 различных химических типов пептидогликана. Большинство различий относится к пептидной части его молекулы.
Две особенности пептидного хвоста заслуживают внимания: наличие аминокислот в D-форме (неприродная конфигурация) и высокое содержание аминокислот с двумя аминогруппами. Это имеет принципиальное значение для пространственной организации пептидогликана. Обе аминогруппы этих аминокислот могут участвовать в образовании пептидных связей, причем вторые аминогруппы — в формировании дополнительных пептидных связей между гетерополимерными цепочками. В большинстве случаев в образовании пептидной связи участвует карбоксильная группа D-аланина одного тетрапептида и свободнаяаминогруппа диаминокислоты другого (рис. 7, А). Иногда связь между тетрапептидами разных гликановых цепей осуществляется с помощью других аминокислот (рис. 7, Б). Нетрудно себе представить, что этим способом можно "сшить" между собой множество гетерополимерных цепей. Частота "сшивок" различна, поскольку не все пептидные хвосты участвуют в формировании межцепочечных связей. Некоторые образуют ковалентные связи с другими химическими молекулами, входящими в состав клеточной стенки, и, наконец, часть тетрапептидных хвостов находится в свободном состоянии.
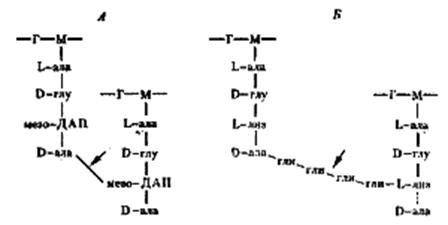
| Рис. 7. Пептидные мостики между гетерополимерными цепочками; Г — N -ацетилглюкозамин: М — N -ацетилмурамовая кислота; ала — аланин; глу — глутаминовая кислота; лиз — лизин; ДАП — диаминопимелиновая кислота; гли — глицин. Стрелками обозначено место действия пенициллина |
Пептидогликан, окружающий протопласт грамположительных эубактерий, — это по существу одна гигантская молекула, "сшитая" с помощью гликозидных и пептидных связей. Именно последние обеспечивают ей трехмерную пространственную организацию.
Кроме пептидогликана в состав клеточных стенок грамположительных эубактерий входит другой уникальный класс химических соединений — тейхоевые кислоты, представляющие собой полимеры, построенные на основе рибита (пятиатомного спирта) или глицерина (трехатомного спирта), остатки которых соединены между собой фосфодиэфирными связями (рис. 8). Некоторые свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками D-аланина, глюкозы, N -ацетилглюкозамина и некоторых других сахаров. Тейхоевые кислоты ковалентно могут соединяться с N -ацетилмурамовой кислотой (см. рис. 6). Поск








