 2018-01-21
2018-01-21 3328
3328Понятие о нейроглии.
Нейроглия – это клетки, окружающие нейроны и входящие вместе с ними в состав ЦНС и ПНС. Количество глиальных клеток на порядок выше количества нервных клеток.
Функции нейроглии:
1. опорная – поддерживает нервные клетки
2. изолирующая – препятствует переходу нервных импульсов с тела одного нейрона на тело другого
3. регуляторная – участвует в регуляции работы ЦНС, в частности, обеспечивая передачу импульсов в нужном направлении
4. трофическая – участвует в обменных процессах нейронов
5. регуляторная – регулирует возбудимость нервных клеток.
(Механизм возникновения потенциала покоя)
Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин:
1) неодинакового распределения ионов по обе стороны мембраны;
2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ.
За счет этих двух факторов создаются условия для движения ионов. Это движение осуществляется без затрат энергии путем пассивного транспорта – диффузией в результате разности концент-рации ионов. Ионы K выходят из клетки и увеличивают положительный заряд на наружной поверхности мембраны, ионы Cl пассивно переходят внутрь клетки, что приводит к увеличению положительного заряда на наружной поверхности клетки. Ионы Na накапливаются на наружной поверхности мембраны и увеличивают ее положительный заряд. Органические соединения остаются внутри клетки. В результате такого движения наружная поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Внутренняя поверхность мембраны может не быть абсолютно отрицательно заряженной, но она всегда заряжена отрицательно по отношению к внешней. Такое состояние клеточной мембраны называется состоянием поляризации. Движение ионов продолжается до тех пор, пока не уравновесится разность потенциалов на мембране, т. е. не наступит электрохимическое равновесие. Момент равновесия зависит от двух сил:
1) силы диффузии;
2) силы электростатического взаимодействия. Значение электрохимического равновесия:
1) поддержание ионной асимметрии;
2) поддержание величины мембранного потенциала на постоянном уровне.
В возникновении мембранного потенциала участвуют сила диффузии (разность концентрации ионов) и сила электростатического взаимодействия, поэтому мембранный потенциал называется концентра-ционно-электрохимическим.
Для поддержания ионной асимметрии электрохимического равновесия недостаточно. В клетке имеется другой механизм – натрий-калиевый насос. Натрий-калиевый насос – механизм обеспечения активного транспорта ионов. В клеточной мембране имеется система переносчиков, каждый из которых связывает три иона Na, которые находятся внутри клетки, и выводит их наружу. С наружной стороны переносчик связывается с двумя ионами K, находящимися вне клетки, и переносит их в цитоплазму. Энергия берется при расщеплении АТФ.
2) (Механизм возникновения потенциала покоя)
Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны.
При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный.
Компоненты потенциала действия:
1) локальный ответ;
2) высоковольтный пиковый потенциал (спайк);
3) следовые колебания.
Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку.
Высоковольтный пиковый потенциал (спайк).
Пик потенциала действия является постоянным компонентом потенциала действия. Он состоит из двух фаз:
1) восходящей части – фазы деполяризации;
2) нисходящей части – фазы реполяризации.
Лавинообразное поступление ионов Na в клетку приводит к изменению потенциала на клеточной мембране. Чем больше ионов Na войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется активационных ворот. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны. Движение ионов Na внутрь клетки продолжается до момента электрохимического равновесия по иону Na Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов Na и от степени проницаемости мембраны к ионам Na. Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. При достижении электрохимического равновесия по ионам Na происходит инактивация активационных ворот, снижается проницаемость к ионам Na и возрастает проницаемость к ионам K. Полного восстановления мембранного потенциала не происходит.
В процессе восстановительных реакций на клеточной мембране регистрируются следовые потенциалы – положительный и отрицательный.
3) (Изменение возбудимости при похождении волны возбуждения)
При развитии потенциала действия происходит изменение возбудимости ткани, причем, это изменение протекает но фазам (рис. 2). Состоянию исходной поляризации мембраны, которую отражает мембранный потенциал покоя, соответствует исходное состояние ее возбудимости и, следовательно, клетки - это нормальный уровень возбудимости. В период предспайка возбудимость ткани повышена, эта фаза возбудимости получила название первичной экзальтации. Во время развития предспайка мембранный потенциал покоя приближается к критическому уровню деполяризации и для достижения последнего достаточна сила раздражителя меньшая, чем пороговая (подпорого-вая).
В период развития спайка (пикового потенциала) идет лавинообразное поступление ионов натрия внутрь клетки, в результате чего происходит перезарядка мембраны и она утрачивает способность отвечать возбуждением на раздражители даже сверхпороговой силы. Эта фаза возбудимости получила название абсолютной рефрактерности (абсолютной невозбудимости). Она длится до конца перезарядки мембраны. Абсолютная рефрактерность, т. е. полная невозбудимость мембраны возникает в связи с тем, что натриевые каналы в начале полностью открываются, а затем инактивируются.
После окончания фазы перезарядки мембраны возбудимость ее постепенно восстанавливается до исходного уровня - фаза относительной рефрактерности. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Так как в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых.
Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости - фаза вторичной экзальтации. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, но сравнению с состоянием покои (исходной поляризацией), то порог раздражения снижен, т. е. возбудимость повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы неполностью. В период развития положительного следового потенциала возбудимость ткани понижена - фаза вторичной рефрактерности. В эту фазу мембранный потенциал увеличивается (состояние гиперполяризации мембраны), удаляясь от критического уровня деполяризации, порог раздражения повышается и новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины. Гиперполяризация мембраны развивается вследствие трех причин: во-первых, продолжающимися выходом ионов калия; во-вторых, открытием, возможно, каналов для хлора и поступление этих ионов в цитоплазму клетки; в-третьих, усиленной работой натрий-калиевого насоса.
4) (Проведение возбуждения по нервным волокнам)
Механизм распространения возбуждения у различных нервных волокон неодинаков. По современным представлениям распространение возбуждения по нервным волокнам осуществляется на основе ионных механизмов генерации потенциала действия.
При распространении возбуждения по безмиелиновому нервному волокну местные электрические токи,, которые возникают между его возбужденным участком, заряженным отрицательно, и невозбужденным, заряженным положительно, вызывают деполяризацию мембраны до критического уровня с последующей генерацией ПД в ближайшей точке невозбужденного участка мембраны. Этот процесс повторяется многократно. На всем протяжении нервного волокна происходит процесс репродукции нового ПД в каждой точке мембраны волокна. Такое проведение возбуждения называете и непрерывным.
Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки (перехватов Ранвье) создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. Местные электрические токи возникают между соседними перехватами Ранвье, т. к. мембрана возбужденного перехвата становится заряженной отрицательно по отношению к поверхности соседнего невозбужденного перехвата. Эти местные токи деполярязуют мембрану невозбужденного перехвата до критического уровня и в нем возникает ПД (рис. 4). Следовательно, возбуждение как бы "перепрыгивает" через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Такой механизм распространения возбуждения называется сальтаторным или скачкообразным. Скорость такого способа проведения возбуждения значительно выше и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов.
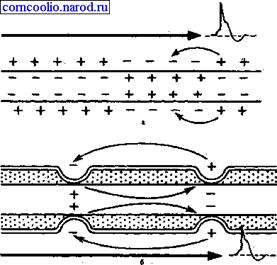
Рис. 4. Схема распространения возбуждения в безмиелиновых (а) и миелиновых (б) нервных волокнах.
"Перепрыгивание" потенциала действия через участок между перехватами возможно потому, что амплитуда ПД в 5-6 раз превышает пороговую величину, необходимую для возбуждения соседнего перехвата. ПД может "перепрыгивать" не только через один, но и через два межперехватных промежутка. Это явление может наблюдаться при снижении возбудимости соседнего перехвата под действием какого-либо фармакологического вещества, например, новокаина, кокаина и др.
Нервные волокна обладают лабильностью - способностью воспроизводить определенное количество циклов возбуждения в единицу времени в соответствии с ритмом действующих раздражителей. Мерой лабильности является максимальное количество циклов возбуждения, которое способно воспроизвести нервное волокно в единицу времени без трансформации ритма раздражения. Лабильность определяется длительностью пика потенциала действия, т. е. фазой абсолютной рефрактерности. Так как длительность абсолютной ре-фрактерности у спайкового потенциала нервного волокна самая короткая, то лабильность его самая высокая. Нервное волокно способно воспроизвести до 1000 импульсов в секунду.
Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента) посредством, например, отравления или повреждения, то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности было названо Н. Е. Введенским парабиозом. В развитии состояния парабиоза можно отметить три, последовательно сменяющих друг друга, фазы: уравнительную, парадоксальную, тормозную.
В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители - больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100 Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т. е. 25 импульсов из 100), то амплитуда ответной реакции становится такой же, как на редкие раздражители (25 Гц) - происходит уравнивание ответной реакции.
В парадоксальную-фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютнойрефрактерности. Следовательно, наблюдается парадокс - на редкие раздражители ответная реакция больше, чем на частые.
В тормозную фазу лабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние.
Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ также связано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам.
Парабиоз - явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности.
Механизм развития парабиотического состояния сводится к следующему. При воздействии на нервное волокно парабиотического фактора нарушается способность мембраны увеличивать натриевую проницаемость в ответ на раздражение. В участке альтерации инак-тивация натриевых каналов, вызванная повреждающим агентом, суммируется с инактивацией, вызываемой нервным импульсом, и возбудимость снижается настолько, что проведение следующего импульса блокируется.
5) (Синапсы, их виды, структурные особенности)
Физиология синапсов.
В ЦНС нервные клетки связаны друг с другом посредством синапсов. Синапс – это структурно функциональное образование, которое обеспечивает передачу возбуждения или торможения с нервного волокна на иннервируемую клетку.
Синапсы по локализации делятся на центральные (расположены в пределах ЦНС, а также в ганглиях вегетативной нервной системы) и периферические (расположены вне ЦНС, обеспечивают связь с клетками иннервируемой ткани).
В функциональном отношении синапсы делятся на возбуждающие, в которых в результате деполяризации постсинаптической мембраны генерируется возбуждающий постсинаптический потенциал, и тормозные, в пресинаптических окончаниях которых выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала.
По механизму передачи синапсы делятся на химические и электрические. Химические синапсы передают возбуждение или торможение за счет особых веществ – медиаторов. В зависимости от вида медиатора химические синапсы подразделяются на:
1. холинергические (медиатор – ацетилхолин)
2. адренергические (медиаторы – адреналин, норадреналин)
По анатомической классификации синапсы делятся на нейросекреторные, нервно-мышечные и межнейронные.
Синапс состоит из трех основных компонентов:
1. пресинаптической мембраны
2. постсинаптической мембраны
3. синаптической щели
Пресинаптическая мембрана является окончанием отростка нервной клетки. Внутри отростка в непосредственной близости от мембраны имеется скопление пузырьков (гранул), содержащих тот или иной медиатор. Пузырьки находятся в постоянном движении.
Постсинаптическая мембрана является частью клеточной мембраны иннервируемой ткани. Постсинаптическая мембрана в отличие от пресинаптической имеет белковые хеморецепторы к биологически активным (медиаторам, гормонам), лекарственным и токсическим веществам. Важная особенность рецепторов постсинаптической мембраны – их химическая специфичность, т.е. способность вступать в биохимическое взаимодействие только с определенным видом медиатора.
Синаптическая щель представляет собой пространство между пре- и постсинаптичекой мембранами, заполненное жидкостью, близкой по составу к плазме крови. Через нее медиатор медленно диффундирует от пресинаптической мембраны к постсинаптической.
Особенности строения нервно-мышечного синапса обусловливают его физиологические свойства.
1. Одностороннее проведение возбуждения (от пре- к постсинаптической мембране), обусловленное наличием чувствительных к медиатору рецепторов только в постсинаптической мембране.
2. Синаптическая задержка проведения возбуждения (время между приходом импульса в пресинаптическое окончание и началом постсинаптического ответа), связанная с малой скоростью диффузии медиатора в синаптическую щель по сравнению со скоростью прохождения импульса по нервному волокну.
3. Низкая лабильность и высокая утомляемость синапса, обусловленная временем распространения предыдущего импульса и наличием у него периода абсолютнойрефрактерности.
4. Высокая избирательная чувствительность синапса к химическим веществам, обусловленная специфичностью хеморецепторов постсинаптической мембраны.
Этапы синаптической передачи.
1. Синтез медиатора. В цитоплазме нейронов и нервных окончаний синтезируются химические медиаторы – биологически активные вещества. Они синтезируются постоянно и депонируются в синаптических пузырьках нервных окончаний.
2. Секреция медиатора. Высвобождение медиатора из синаптических пузырьков имеет квантовый характер. В состоянии покоя оно незначительно, а под влиянием нервного импульса резко усиливается.
3. Взаимодействие медиатора с рецепторами постсинаптической мембраны. Это взаимодействие заключается в избирательном изменении проницаемости ионоселективных каналов эффекторной клетки в области активных центров связывания с медиатором. Взаимодействие медиатора со своими рецепторами может вызвать возбуждение или торможение нейрона, сокращение мышечной клетки, образование и выделение гормонов секреторными клетками. В случае увеличения проницаемости натриевых и кальциевых каналов усиливается поступление Na и Ca в клетку с последующей деполяризацией мембраны, возникновением ПД и дальнейшей передачей нервного импульса. Такие синапсы называются возбуждающими. Если повышается проницаемость калиевых каналов и каналов для хлора, наблюдается избыточный выход К из клетки с одновременной диффузией в нее Cl, что приводит к гиперполяризации мембраны, снижению ее возбудимости и развитию тормозных постсинаптических потенциалов. Передача нервных импульсов затрудняется или совсем прекращается. Такие синапсы называются тормозными.
Рецепторы, взаимодействующие с АХ, называются холинорецепторами. В функциональном отношении они разделяются на две группы: М - и Н-холинорецепторы. В синапсах скелетных мышц присутствуют только Н-холинорецепторы, тогда как в мышцах внутренних органов – преимущественно М-холинорецепторы.
Рецепторы, взаимодействующие с НА, называются адренорецепторами. В функциональном отношении они делятся на альфа- и бета-адренорецепторы. В постсинаптической мембране гладкомышечных клеток внутренних органов и кровеносных сосудов часто соседствуют оба вида адренорецепторов. Действие НА является деполяризующим, если он взаимодействует с альфа-адренорецепторами (сокращение мышечной оболочки стенок кровеносных сосудов или кишечника), или тормозным – при взаимодействии с бета-адренорецепторами (их расслабление).
4. Инактивация медиатора. Инактивация (полная потеря активности) медиатора необходима для реполяризации постсинаптической мембраны и восстановления исходного уровня МП. Наиболее важным путем инактивации является гидролитическое расщепление с помощью ингибиторов. Для АХ ингибитором является холинэстераза, для НА и адреналина – моноаминоксидаза и катехолоксиметилтрансфераза.
Другой путь удаления медиатора из синаптической щели – «обратный захват» его пресинаптическими окончаниями (пиноцитоз) и обратный аксонный транспорт, особенно выраженный для катехоламинов.
В основе координационной деятельности ЦНС лежит взаимодействие процессов возбуждения и торможения.
Возбуждение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся повышением функций ткани.
Торможение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся снижением функций ткани.
Первичное торможение в ЦНС возникает за счет тормозных нейронов. Это особый вид вставочных нейронов, которые при передаче импульса выделяют тормозной медиатор. Различают 2 вида первичного торможения: постсинаптическое и пресинаптическое.
Постсинаптическое торможение возникает, если аксон тормозного нейрона образует синапс с телом нейрона и, выделяя медиатор, вызывает гиперполяризацию клеточной мембраны, тормозя активность клетки.
Пресинаптическое торможение возникает, когда аксон тормозного нейрона образует синапс с аксоном возбуждающего нейрона, препятствуя проведению импульса.
6) (Спинной мозг, его функции, участие в регуляции мышечного тонуса)
Спинной мозг выполняет рефлекторную и проводниковую функции. Первая обеспечивается его нервными центрами, вторая проводящими путями.
Он имеет сегментарное строение. Причем деление на сегменты является функциональным. Каждый сегмент образует передние и задние корешки. Задние являются чувствительными, т.е. афферентными, передние двигательными, эфферентными. Эта закономерность называется законом Белла-Мажанди. Корешки каждого сегмента иннервируют 3 метамера тела, но в результате перекрывания каждый метамер иннервируется тремя сегментами. Поэтому при поражении передних корешков одного сегмента, двигательная активность соответствующего метамера лишь ослабляется.
Морфологически тела нейронов спинного мозга образуют его серое вещество. Функционально все его нейроны делятся на мотонейроны, вставочные, нейроны симпатического и парасимпатического отделов вегетативной нервной системы. Мотонейроны, в зависимости от функционального значения, разделяются на альфа – и гамма-мотонейроны. К a-мотонейронам идут волокна афферентных путей, которые начинаются от интрафузальных, т.е. рецепторных мышечных клеток. Тела a-мотонейронов расположены в передних рогах спинного мозга, а их аксоны иннервируют скелетные мышцы. Гамма-мотонейроны регулируют напряжение мышечных веретен т.е. интрафузальных волокон. Таким образом они участвуют в регуляции сокращений скелетных мышц. Поэтому при перерезке передних корешков мышечный тонус исчезает.
Интернейроны обеспечивают связь между центрами спинного мозга и вышележащих отделов ЦНС.
Нейроны симпатического отдела вегетативной нервной системы находятся в боковых рогах грудных сегментов, а парасимпатического в крестцовом отделе.
Проводниковая функция состоит в обеспечении связи периферических рецепторов, центров спинного мозга с вышележащими отделами ЦНС, а также его нервных центров между собой. Она осуществляется проводящими путями. Все пути спинного мозга делятся на собственные или проприоспинальные, восходящие и нисходящие. Проприоспинальные пути связывают между собой нервные центры разных сегментов спинного мозга. Их функция заключается в координации тонуса мышц, движений различных метамеров туловища.
К восходящим путям относятся несколько трактов. Пучки Голля и Бурдаха проводят нервные импульсы от проприорецепторов мышц и сухожилий к соответствующим ядрам продолговатого мозга, а затем таламусу и соматосенсорным зонам коры. Благодаря этим путям производится оценка и коррекция позы туловища. Пучки Говерса и Флексига передают возбуждение от проприорецепторов, механорецепторов кожи к мозжечку. За счет этого обеспечивается восприятие и бессознательная координация позы. Спиноталамические тракты проводят сигналы от болевых, температурных, тактильных рецепторов кожи к таламусу, а затем соматосенсорные зоны коры. Они обеспечивают восприятие соответствующих сигналов и формирование чувствительности.
Нисходящие пути также образованы несколькими трактами. Кортикоспинальные пути идут от пирамидных и экстрапирамидных нейронов коры к a-мотонейронам спинного мозга. За счет них осуществляется регуляция произвольных движений. Руброспинальный путь проводит сигналы от красного ядра среднего мозга к a-мотонейронам мышц сгибателей. Вестибулоспинальный путь передает сигналы от вестибулярных ядер продолговатого мозга, в первую очередь ядра Дейтерса, к a-мотонейронам мышц разгибателей. За счет этих двух путей регулируется тонус соответствующих мышц при изменениях положения тела.
Все рефлексы спинного мозга делятся на соматические, т.е. двигательные и вегетативные. Соматические рефлексы делятся на сухожильные или миотатические и кожные. Сухожильные рефлексы возникают при механическом раздражении мышц и сухожилий. Их небольшое растяжение приводит к возбуждению сухожильных рецепторов и a-мотонейронов спинного мозга. В результате возникает сокращение мышц, в первую очередь разгибателей. К сухожильным рефлексам относятся коленный, ахиллов, локтевой, кистевой и др., возникающие при механическом раздражении соответствующих сухожилий. Например, коленный является простейшим моносинаптическим, так как в его центральной части только один синапс. Кожные рефлексы обусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной (объяснение). Спинальные нервные центры находятся под контролем вышележащих. Поэтому после перерезки между продолговатым и спинным мозгом возникает спинальный шок и тонус всех мышц значительно уменьшится.
Вегетативные рефлексы спинного мозга делятся на симпатические и парасимпатические. Те и другие проявляются реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны спинного мозга образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыведения, дефекации, эрекции, эйякуляции и т.д.
7) (Продолговатый мозг и мост, их функции, участие в регуляции мышечного тонуса)
Продолговатый мозг
Особенности функциональной организации. Продолговатый мозг (medullaoblongata) у человека имеет длину около 25 мм. Он является продолжением спинного мозга. Структурно по разнообразию и строению ядер продолговатый мозг сложнее, чем спинной. В отличие от спинного мозга он не имеет метамерного, повторяемого строения, серое вещество в нем расположено не в центре, а ядрами к периферии.
В продолговатом мозге находятся оливы, связанные со спинным мозгом, экстрапирамидной системой и мозжечком — это тонкое и клиновидное ядра проприоцептивной чувствительности (ядра Голля и Бурдаха). Здесь же находятся перекресты нисходящих пирамидных путей и восходящих путей, образованных тонким и клиновидным пучками (Голля и Бурдаха), ретикулярная формация.
Продолговатый мозг за счет своих ядерных образований и ретикулярной формации участвует в реализации вегетативных, соматических, вкусовых, слуховых, вестибулярных рефлексов. Особенностью продолговатого мозга является то, что его ядра, возбуждаясь последовательно, обеспечивают выполнение сложных рефлексов, требующих последовательного включения разных мышечных групп, что наблюдается, например, при глотании.
В продолговатом мозге расположены ядра следующих черепных нервов:
пара VIII черепных нервов — преддверно-улитковый нерв состоит из улитковой и преддверной частей. Улитковое ядро лежит в продолговатом мозге;
пара IX — языкоглоточный нерв (п. glossopharyngeus); его ядро образовано 3 частями — двигательной, чувствительной и вегетативной. Двигательная часть участвует в иннервации мышц глотки и полости рта, чувствительная — получает информацию от рецепторов вкуса задней трети языка; вегетативная иннервирует слюнные железы;
пара X — блуждающий нерв (n.vagus) имеет 3 ядра: вегетативное иннервирует гортань, пищевод, сердце, желудок, кишечник, пищеварительные железы; чувствительное получает информацию от рецепторов альвеол легких и других внутренних органов и двигательное (так называемое обоюдное) обеспечивает последовательность сокращения мышц глотки, гортани при глотании;
пара XI — добавочный нерв (n.accessorius); его ядро частично расположено в продолговатом мозге;
пара XII — подъязычный нерв (n.hypoglossus) является двигательным нервом языка, его ядро большей частью расположено в продолговатом мозге.
Сенсорные функции. Продолговатый мозг регулирует ряд сенсорных функций: рецепцию кожной чувствительности лица — в сенсорном ядре тройничного нерва; первичный анализ рецепции вкуса — в ядре языкоглоточного нерва; рецепцию слуховых раздражений — в ядре улиткового нерва; рецепцию вестибулярных раздражений — в верхнем вестибулярном ядре. В задневерхних отделах продолговатого мозга проходят пути кожной, глубокой, висцеральной чувствительности, часть из которых переключается здесь на второй нейрон (тонкое и клиновидное ядра). На уровне продолготоватого мозга перечисленные сенсорные функции реализуют первичный анализ силы и качества раздражения, далее обработанная информация передается в подкорковые структуры для определения биологической значимости данного раздражения.
Проводниковые функции. Через продолготоватый мозг проходят все восходящие и нисходящие пути спинного мозга: спинно-таламический, кортикоспинальный, руброспинальный. В нем берут начало вестибулоспинальный, оливоспинальный и ретикулоспинальный тракты, обеспечивающие тонус и координацию мышечных реакций. В продолговатом мозге заканчиваются пути из коры большого мозга — корковоретикулярные пути. Здесь заканчиваются восходящие пути проприоцептивной чувствительности из спинного мозга: тонкого и клиновидного. Такие образования головного мозга, как мост, средний мозг, мозжечок, таламус, гипоталамус и кора большого мозга, имеют двусторонние связи с продолговатым мозгом. Наличие этих связей свидетельствует об участии продолговатого мозга в регуляции тонуса скелетной мускулатуры, вегетативных и высших интегративных функций, анализе сенсорных раздражений.
Рефлекторные функции. Многочисленные рефлексы продолговатого мозга делят на жизненно важные и нежизненно важные, однако такое представление достаточно условно. Дыхательные и сосудодвигательные центры продолговатого мозга можно отнести к жизненно важным центрам, так как в них замыкается ряд сердечных и дыхательных рефлексов.
Продолговатый мозг организует и реализует ряд защитных рефлексов: рвоты, чиханья, кашля, слезоотделения, смыкания век. Эти рефлексы реализуются благодаря тому, что информация о раздражении рецепторов слизистой оболочки глаза, полости рта, гортани, носоглотки через чувствительные ветви тройничного и языкоглоточного нервов попадает в ядра продолговатого мозга, отсюда идет команда к двигательным ядрам тройничного, блуждающего, лицевого, языкоглоточного, добавочного или подъязычного нервов, в результате реализуется тот или иной защитный рефлекс. Точно так же за счет последовательного включения мышечных групп головы, шеи, грудной клетки и диафрагмы организуются рефлексы пищевого поведения: сосания, жевания, глотания.
Кроме того, продолговатый мозг организует рефлексы поддержания позы. Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов в верхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейронов медиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо.
Изменение позы осуществляется за счет статических и статокинетических рефлексов. Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела. Статокинетические рефлексы продолговатого мозга обеспечивают перераспределение тонуса мышц туловища для организации позы, соответствующей моменту прямолинейного или вращательного движения.
Большая часть автономных рефлексов продолговатого мозга реализуется через расположенные в нем ядра блуждающего нерва, которые получают информацию о состоянии деятельности сердца, сосудов, пищеварительного тракта, легких, пищеварительных желез и др. В ответ на эту информацию ядра организуют двигательную и секреторную реакции названных органов.
Возбуждение ядер блуждающего нерва вызывает усиление сокращения гладких мышц желудка, кишечника, желчного пузыря и одновременно расслабление сфинктеров этих органов. При этом замедляется и ослабляется работа сердца, сужается просвет бронхов.
Деятельность ядер блуждающего нерва проявляется также в усилении секреции бронхиальных, желудочных, кишечных желез, в возбуждении поджелудочной железы, секреторных клеток печени.
В продолговатом мозге локализуется центр слюноотделения, парасимпатическая часть которого обеспечивает усиление общей секреции, а симпатическая — белковой секреции слюнных желез.
В структуре ретикулярной формации продолговатого мозга расположены дыхательный и сосудодвигательный центры. Особенность этих центров в том, что их нейроны способны возбуждаться рефлекторно и под действием химических раздражителей.
Дыхательный центр локализуется в медиальной части ретикулярной формации каждой симметричной половины продолговатого мозга и разделен на две части, вдоха и выдоха.
В ретикулярной формации продолговатого мозга представлен другой жизненно важный центр — сосудодвигательный центр (регуляции сосудистого тонуса). Он функционирует совместно с вышележащими структурами мозга и прежде всего с гипоталамусом. Возбуждение сосудодвигательного центра всегда изменяет ритм дыхания, тонус бронхов, мышц кишечника, мочевого пузыря, цилиарной мышцы и др. Это обусловлено тем, что ретикулярная формация продолговатого мозга имеет синаптические связи с гипоталамусом и другими центрами.
В средних отделах ретикулярной формации находятся нейроны, образующие ретикулоспинальный путь, оказывающий тормозное влияние на мотонейроны спинного мозга. На дне IV желудочка расположены нейроны «голубого пятна». Их медиатором является норадреналин. Эти нейроны вызывают активацию ретикулоспинального пути в фазу «быстрого» сна, что приводит к торможению спинальных рефлексов и снижению мышечного тонуса.
Симптомы повреждения. Повреждение левой или правой половины продолговато мозга выше перекреста восходящих путей проприоцептивной чувствительности вызывает на стороне повреждения нарушения чувствительности и работы мышц лица и головы. В то же время на противоположной стороне относительно стороны повреждения наблюдаются нарушения кожной чувствительности и двигательные параличи туловища и конечностей. Это объясняется тем, что восходящие и нисходящие проводящие пути из спинного мозга и в спинной мозг перекрещиваются, а ядра черепных нервов иннервируют свою половину головы, т. е. черепные нервы не перекрещиваются.
Мост
Мост (ponscerebri, ponsVarolii) располагается выше продолговатого мозга и выполняет сенсорные, проводниковые, двигательные, интегративные рефлекторные функции.
В состав моста входят ядра лицевого, тройничного, отводящего, преддверно-улиткового нерва (вестибулярные и улитковые ядра), ядра преддверной части преддверно-улиткового нерва (вестибулярного нерва): латеральное (Дейтерса) и верхнее (Бехтерева). Ретикулярная формация моста тесно связана с ретикулярной формацией среднего и продолговатого мозга.
Важной структурой моста является средняя ножка мозжечка. Именно она обеспечивает функциональные компенсаторные и морфологические связи коры большого мозга с полушариями мозжечка.
Сенсорные функции моста обеспечиваются ядрами преддверно-улиткового, тройничного нервов. Улитковая часть преддверно-улиткового нерва заканчивается в мозге в улитковых ядрах; преддверная часть преддверно-улиткового нерва — в треугольном ядре, ядре Дейтерса, ядре Бехтерева. Здесь происходит первичный анализ вестибулярных раздражений их силы и направленности.
Чувствительное ядро тройничного нерва получает сигналы от рецепторов кожи лица, передних отделов волосистой части головы, слизистой оболочки носа и рта, зубов и конъюнктивы глазного яблока. Лицевой нерв (п. facialis) иннервирует все мимические мышцы лица. Отводящий нерв (п. abducens) иннервирует прямую латеральную мышцу, отводящую глазное яблоко кнаружи.
Двигательная порция ядра тройничного нерва (п. trigeminus) иннервирует жевательные мышцы, мышцу, натягивающую барабанную перепонку, и мышцу, натягивающую небную занавеску.
Проводящая функция моста. Обеспечивается продольно и поперечно расположенными волокнами. Поперечно расположенные волокна образуют верхний и нижний слои, а между ними проходят идущие из коры большого мозга пирамидные пути. Между поперечными волокнами расположены нейронные скопления — ядра моста. От их нейронов начинаются поперечные волокна, которые идут на противоположную сторону моста, образуя среднюю ножку мозжечка и заканчиваясь в его коре.
В покрышке моста располагаются продольно идущие пучки волокон медиальной петли (lemniscusmedialis). Они пересекаются поперечно идущими волокнами трапециевидного тела (corpustrapezoideum), представляющие собой аксоны улитковой части преддверно-улиткового нерва противоположной стороны, которые заканчиваются в ядре верхней оливы (olivasuperior). От этого ядра идут пути боковой петли (lemniscuslateralis), которые направляются в заднее четверохолмие среднего мозга и в медиальные коленчатые тела промежуточного мозга.
В покрышке мозга локализуются переднее и заднее ядра трапециевидного тела и латеральной петли. Эти ядра вместе с верхней оливой обеспечивают первичный анализ информации от органа слуха и затем передают информацию в задние бугры четверохолмий.
В покрышке также расположены длинный медиальный и тектоспинальный пути.
Собственные нейроны структуры моста образуют его ретикулярную формацию, ядра лицевого, отводящего нервов, двигательной порции ядра и среднее сенсорное ядро тройничного нерва.
Ретикулярная формация моста является продолжением ретикулярной формации продолговатого мозга и началом этой же системы среднего мозга. Аксоны нейронов ретикулярной формации моста идут в мозжечок, в спинной мозг (ретикулоспинальный путь). Последние активируют нейроны спинного мозга.
Ретикулярная формация моста влияет на кору большого мозга, вызывая ее пробуждение или сонное состояние. В ретикулярной формации моста находятся две группы ядер, которые относятся к общему дыхательному центру. Один центр активирует центр вдоха продолговатого мозга, другой — центр выдоха. Нейроны дыхательного центра, расположенные в мосте, адаптируют работу дыхательных клеток продолговатого мозга в соответствии с меняющимся состоянием организма.
8) (Средний мозг, его функции, участие в регуляции мышечного тонуса)
Морфофункциональная организация. Средний мозг (mesencephalon) представлен четверохолмием и ножками мозга. Наиболее крупными ядрами среднего мозга являются красное ядро, черное вещество и ядра черепных (глазодвигательного и блокового) нервов, а также ядра ретикулярной формации.
Сенсорные функции. Реализуются за счет поступления в него зрительной, слуховой информации.
Проводниковая функция. Заключается в том, что через него проходят все восходящие пути к вышележащим таламусу (медиальная петля, спииноталамический путь), большому мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу. Это пирамидный путь, корково-мостовые волокна, руброретикулоспинальный путь.
Двигательная функция. Реализуется за счет ядра блокового нерва (n. trochlearis), ядер глазодвигательного нерва (п. oculomotorius), красного ядра (nucleusruber), черного вещества (substantianigra).
Красные ядра располагаются в верхней части ножек мозга. Они связаны с корой большого мозга (нисходящие от коры пути), подкорковыми ядрами, мозжечком, спинным мозгом (красноядерно-спинномозговой путь). Базальные ганглии головного мозга, мозжечок имеют свои окончания в красных ядрах. Нарушение связей красных ядер с ретикулярной формацией продолговатого мозга ведет к децеребрационной ригидности. Это состояние характеризуется сильным напряжением мышц-разгибателей конечностей, шеи, спины. Основной причиной возникновения децеребрационной ригидности служит выраженное активирующее влияние латерального вестибулярного ядра (ядро Дейтерса) на мотонейроны разгибателей. Это влияние максимально в отсутствие тормозных влияний красного ядра и вышележащих структур, а также мозжечка. При перерезке мозга ниже ядра латерального вестибулярного нерва децеребрационная ригидность исчезает.
Красные ядра, получая информацию от двигательной зоны коры большого мозга, подкорковых ядер и мозжечка о готовящемся движении и состоянии опорно-двигательного аппарата, посылают корригирующие импульсы к мотонейронам спинного мозга по руброспинальному тракту и тем самым регулируют тонус мускулатуры, подготавливая его уровень к намечающемуся произвольному движению.
Другое функционально важное ядро среднего мозга — черное вещество — располагается в ножках мозга, регулирует акты жевания, глотания (их последовательность), обеспечивает точные движения пальцев кисти руки, например при письме. Нейроны этого ядра способны синтезировать медиатор дофамин, который поставляется аксональным транспортом к базальным ганглиям головного мозга. Поражение черного вещества приводит к нарушению пластического тонуса мышц. Тонкая регуляция пластического тонуса при игре на скрипке, письме, выполнении графических работ обеспечивается черным веществом. В то же время при длительном удержании определенной позы происходят пластические изменения в мышцах за счет изменения их коллоидных свойств, что обеспечивает наименьшие затраты энергии. Регуляция этого процесса осуществляется клетками черного вещества.
Нейроны ядер глазодвигательного и блокового нервов регулируют движение глаза вверх, вниз, наружу, к носу и вниз к углу носа. Нейроны добавочного ядра глазодвигательного нерва (ядро Якубовича) регулируют просвет зрачка и кривизну хрусталика.
Рефлекторные функции. Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Верхние из них являются первичными подкорковыми центрами зрительного анализатора (вместе с латеральными коленчатыми телами промежуточного мозга), нижние — слухового (вместе с медиальными коленчатыми телами промежуточного мозга). В них происходит первичное переключение зрительной и слуховой информации. От бугров четверохолмия аксоны их нейронов идут к ретикулярной формации ствола, мотонейронам спинного мозга. Нейроны четверохолмия могут быть полимодальными и детекторными. В последнем случае они реагируют только на один признак раздражения, например смену света и темноты, направление движения светового источника и т. д. Основная функция бугров четверохолмия — организация реакции настораживания и так называемых старт-рефлексов на внезапные, еще не распознанные, зрительные или звуковые сигналы. Активация среднего мозга в этих случаях через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию, к оборонительной реакции.
Четверохолмие организует ориентировочные зрительные и слуховые рефлексы.
У человека четверохолмный рефлекс является сторожевым. В случаях повышенной возбудимости четверохолмий при внезапном звуковом или световом раздражении у человека возникает вздрагивание, иногда вскакивание на ноги, вскрикивание, максимально быстрое удаление от раздражителя, подчас безудержное бегство.
При нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое. Следовательно, четверохолмия принимают участие в организации произвольных движений.
Ретикулярная формация ствола мозга
Ретикулярная формация (formatioreticularis; РФ) мозга представлена сетью нейронов с многочисленными диффузными связями между собой и практически со всеми структурами центральной нервной системы. РФ располагается в толще серого вещества продолговатого, среднего, промежуточного мозга и изначально связана с РФ спинного мозга. В связи с этим целесообразно ее рассмотреть как единую систему. Сетевые связи нейронов РФ между собой позволили Дейтерсу назвать ее ретикулярной формацией мозга.
РФ имеет прямые и обратные связи с корой большого мозга, базальными ганглиями, промежуточным мозгом, мозжечком, средним, продолговатым и спинным мозгом.
Основной функцией РФ является регуляция уровня активности коры большого мозга, мозжечка, таламуса, спинного мозга.
С одной стороны, генерализованный характер влияния РФ на многие структуры мозга дал основание считать ее неспецифической системой. Однако исследования с раздражением РФ ствола показали, что она может избирательно оказывать активирующее или тормозящее влияние на разные формы поведения, на сенсорные, моторные, висцеральные системы мозга. Сетевое строение обеспечивает высокую надежность функционирования РФ, устойчивость к повреждающим воздействиям, так как локальные повреждения всегда компенсируются за счет сохранившихся элементов сети. С другой стороны, высокая надежность функционирования РФ обеспечивается тем, что раздражение любой из ее частей отражается на активности всей РФ данной структуры за счет диффузности связей.
Большинство нейронов РФ имеет длинные дендриты и короткий аксон. Существуют гигантские нейроны с длинным аксоном, образующие пути из РФ в другие области мозга, например в нисходящем направлении, ретикулоспинальный и руброспинальный. Аксоны нейронов РФ образуют большое число коллатералей и синапсов, которые оканчиваются на нейронах различных отделов мозга. Аксоны нейронов РФ, идущие в кору большого мозга, заканчиваются здесь на дендритах I и II слоев.
Активность нейронов РФ различна и в принципе сходна с активностью нейронов других структур мозга, но среди нейронов РФ имеются такие, которые обладают устойчивой ритмической активностью, не зависящей от приходящих сигналов.
В то же время в РФ среднего мозга и моста имеются нейроны, которые в покое «молчат», т. е. не генерируют импульсы, но возбуждаются при стимуляции зрительных или слуховых рецепторов. Это так называемые специфические нейроны, обеспечивающие быструю реакцию на внезапные, неопознанные сигналы. Значительное число нейронов РФ являются полисенсорными.
В РФ продолговатого, среднего мозга и моста конвергируют сигналы различной сенсорности. На нейроны моста приходят сигналы преимущественно от соматосенсорных систем. Сигналы от зрительной и слуховой сенсорных систем в основном приходят на нейроны РФ среднего мозга.
РФ контролирует передачу сенсорной информации, идущей через ядра таламуса, за счет того, что при интенсивном внешнем раздражении нейроны неспецифических ядер таламуса затормаживаются, тем самым снимается их тормозящее влияние с релейных ядер того же таламуса и облегчается передача сенсорной информации в кору большого мозга.
В РФ моста, продолговатого, среднего мозга имеются нейроны, которые реагируют на болевые раздражения, идущие от мышц или внутренних органов, что создает общее диффузное дискомфортное, не всегда четко локализуемое, болевое ощущение «тупой боли».
Повторение любого вида стимуляции приводит к снижению импульсной активности нейронов РФ, т. е. процессы адаптации (привыкания) присущи и нейронам РФ ствола мозга.
РФ ствола мозга имеет прямое отношение к регуляции мышечного тонуса, поскольку на РФ ствола мозга поступают сигналы от зрительного и вестибулярного анализаторов и мозжечка. От РФ к мотонейронам спинного мозга и ядер черепных нервов поступают сигналы, организующие положение головы, туловища и т. д.
Ретикулярные пути, облегчающие активность моторных систем спинного мозга, берут начало от всех отделов РФ. Пути, идущие от моста, тормозят активность мотонейронов спинного мозга, иннервирующих мышцы-сгибатели, и активируют мотонейроны мышц-разгибателей. Пути, идущие от РФ продолговатого мозга, вызывают противоположные эффекты. Раздражение РФ приводит к тремору, повышению тонуса мышц. После прекращения раздражения вызванный им эффект сохраняется длительно, видимо, за счет циркуляции возбуждения в сети нейронов.
РФ ствола мозга участвует в передаче информации от коры большого мозга, спинного мозга к мозжечку и, наоборот, от мозжечка к этим же системам. Функция данных связей заключается в подготовке и реализации моторики, связанной с привыканием, ориентировочными реакциями, болевыми реакциями, организацией ходьбы, движениями глаз.
Регуляция вегетативной деятельности организма РФ описана в разделе 4.3, здесь же заметим, что наиболее четко эта регуляция проявляется в функционировании дыхательного и сердечно-сосудистых центров. В регуляции вегетативных функций большое значение имеют так называемые стартовые нейроны РФ. Они дают начало циркуляции возбуждения внутри группы нейронов, обеспечивая тонус регулируемых вегетативных систем.
Влияния РФ можно разделить в целом на нисходящие и восходящие. В свою очередь каждое из этих влияний имеет тормозное и возбуждающее действие.
Восходящие влияния РФ на кору большого мозга повышают ее тонус, регулируют возбудимость ее нейронов, не изменяя специфику ответов на адекватные раздражения. РФ влияет на функциональное состояние всех сенсорных областей мозга, следовательно, она имеет значение в интеграции сенсорной информации от разных анализаторов.
РФ имеет прямое отношение к регуляции цикла бодрствование—сон. Стимуляция одних структур РФ приводит к развитию сна, стимуляция других вызывает пробуждение. Г. Мэгун и Д. Моруцци выдвинули концепцию, согласно которой все виды сигналов, идущих от периферических рецепторов, достигают по коллатералям РФ продолговатого мозга и моста, где переключаются на нейроны, дающие восходящие пути в таламус и затем в кору большого мозга.
Возбуждение РФ продолговатого мозга или моста вызывает синхронизацию активности коры большого мозга, появление медленных ритмов в ее электрических показателях, сонное торможение.
Возбуждение РФ среднего мозга вызывает противоположный эффект пробуждения: десинхронизацию электрической активности коры, появление быстрых низкоамплитудных β-подобных ритмов в электроэнцефалограмме.
Г. Бремер (1935) показал, что если перерезать мозг между передними и задними буграми четверохолмия, то животное перестает реагировать на все виды сигналов; если же перерезку произвести между продолговатым и средним мозгом (при этом РФ сохраняет связь с передним мозгом), то животное реагирует на свет, звук и другие сигналы. Следовательно, поддержание активного анализирующего состояния мозга возможно при сохранении связи с передним мозгом.
Реакция активации коры большого мозга наблюдается при раздражении РФ продолговатого, среднего, промежуточного мозга. В то же время раздражение некоторых ядер таламуса приводит к возникновению ограниченных локальных участков возбуждения, а не к общему ее возбуждению, как это бывает при раздражении других отделов РФ.
РФ ствола мозга может оказывать не только возбуждающее, но и тормозное влияние на активность коры мозга.
Нисходящие влияния РФ ствола мозга на регуляторную деятельность спинного мозга были установлены еще И. М. Сеченовым (1862). Им было показано, что при раздражении среднего мозга кристалликами соли у лягушки рефлексы отдергивания лапки возникают медленно, требуют более сильного раздражения или не появляются вообще, т. е. тормозятся.
Г. Мэгун (1945—1950), нанося локальные раздражения на РФ продолговатого мозга, нашел, что при раздражении одних точек тормозятся, становятся вялыми рефлексы сгибания передней лапы, коленный, роговичный. При раздражении РФ в других точках продолговатого мозга эти же рефлексы вызывались легче, были сильнее, т. е. их реализация облегчалась. По мнению Мэгуна, тормозные влияния на рефлексы спинного мозга может оказывать только РФ продолговатого мозга, а облегчающие влияния регулируются всей РФ ствола и спинного мозга.
9) (Мозжечок, его участие в регуляции моторных и вегетативных функций)
Мозжечок (cerebellum, малый мозг) — одна из интегративных структур головного мозга, принимающая участие в координации и регуляции произвольных, непроизвольных движений, в регуляции вегетативных и поведенческих функций.
Особенности морфофункциональной организации и связи мозжечка. Реализация указанных функций обеспечивается следующими морфологическими особенностями мозжечка:
1) кора мозжечка построена достаточно однотипно, имеет стереотипные связи, что создает условия для быстрой обработки информации;
2) основной нейронный элемент коры — клетка Пуркинье, имеет большое количество входов и формирует единственный аксонный выход из мозжечка, коллатерали которого заканчиваются на ядерных его структурах;
3) на клетки Пуркинье проецируются практически все видысенсорных раздражений: проприоцептивные, кожные, зрительные,слуховые, вестибулярные и др.;
4) выходы из мозжечка обеспечивают его связи с корой большого мозга, со стволовыми образованиями и спинным мозгом.
Мозжечок анатомически и функционально делится на старую, древнюю и новую части.
К старой части мозжечка (archicerebellum) — вестибулярный мозжечок — относится клочково-флоккулярная доля. Эта часть имеет наиболее выраженные связи с вестибулярным анализатором, что объясняет значение мозжечка в регуляции равновесия.
Древняя часть мозжечка (paleocerebellum) — спинальный мозжечок — состоит из участков червя и пирамиды мозжечка, язычка, околоклочкового отдела и получает информацию преимущественно от проприорецептивных систем мышц, сухожилий, надкостницы, оболочек суставов.
Новый мозжечок (neocerebellum) включает в себя кору полушарий мозжечка и участки червя; он получает информацию от коры, преимущественно по лобно-мостомозжечковому пути, от зрительных и слуховых рецептирующих систем, что свидетельствует об его участии в анализе зрительных, слуховых сигналов и организации на них реакции.
Кора мозжечка имеет специфическое, нигде в ЦНС не повторяющееся, строение. Верхний (I) слой коры мозжечка — молекулярный слой, состоит из параллельных волокон, разветвлений дендритов и аксонов II и III слоев. В нижней части молекулярного слоя встречаются корзинчатые и звездчатые клетки, которые обеспечивают взаимодействие клеток Пуркинье.
Средний (II) слой коры образован клетками Пуркинье, выстроенными в один ряд и имеющими самую мощную в ЦНС дендритную систему. На дендритном поле одной клетки Пуркинье может быть до 60 000 синапсов. Следовательно, эти клетки выполняют задачу сбора, обработки и передачи информации. Аксоны клеток Пуркинье являются единственным путем, с помощью которого кора мозжечка передает информацию в его ядра и ядра структуры большого мозга.
Под II слоем коры (под клетками Пуркинье) лежит гранулярный (III) слой, состоящий из клеток-зерен, число которых достигает 10 млрд. Аксоны этих клеток поднимаются вверх, Т-образно делятся на поверхности коры, образуя дорожки контактов с клетками Пуркинье. Здесь же лежат клетки Гольджи.
Из мозжечка информация уходит через верхние и нижние ножки. Через верхние ножки сигналы идут в таламус, в мост, красное ядро, ядра ствола мозга, в ретикулярную формацию среднего мозга. Через нижние ножки мозжечка сигналы идут в продолговатый мозг к его вестибулярным ядрам, оливам, ретикулярной формации. Средние ножки мозжечка связывают новый мозжечок с лобной долей мозга.
Импульсная активность нейронов регистрируется в слое клеток Пуркинье и гранулярном слое, причем частота генерации импульсов этих клеток колеблется от 20 до 200 в секунду. Клетки ядер мозжечка генерируют импульсы значительно реже — 1—3 импульса в секунду.
Стимуляция верхнего слоя коры мозжечка приводит к длительному (до 200 мс) торможению активности клеток Пуркинье. Такое же их торможение возникает при световых и звуковых сигналах. Суммарные изменения электрической активности коры мозжечка на раздражение чувствительного нерва любой мышцы выглядят в форме позитивного колебания (торможение активности коры, гиперполяризация клеток Пуркинье), которое наступает через 15— 20 мс и длится 20—30 мс, после чего возникает волна возбуждения, длящаяся до 500 мс (деполяризация клеток Пуркинье).
В кору мозжечка от кожных рецепторов, мышц, суставных оболочек, надкостницы сигналы поступают по так называемым спинно-мозжечковым трактам: по заднему (дорсальному) и переднему (вентральному). Эти пути к мозжечку проходят через нижнюю оливу продолговатого мозга. От клеток олив идут так называемые лазающие волокна, которые ветвятся на дендритах клеток Пуркинье.
Ядра моста посылают афферентные пути в мозжечок, образующие мшистые волокна, которые оканчиваются на клетках-зернах III слоя коры мозжечка. Между мозжечком и голубоватым местом среднего мозга существует афферентная связь с помощью адренергических волокон. Эти волокна способны диффузно выбрасывать норадреналин в межклеточное пространство коры мозжечка, тем самым гуморально изменяют состояние возбудимости его клеток.
Аксоны клеток III слоя коры мозжечка вызывают торможение клеток Пуркинье и клеток-зерен своего же слоя.
Клетки Пуркинье в свою очередь тормозят активность нейронов ядер мозжечка. Ядра мозжечка имеют высокую тоническую активность и регулируют тонус ряда моторных центров промежуточного, среднего, продолговатого, спинного мозга.
Подкорковая система мозжечка состоит из трех функционально разных ядерных образований: ядра шатра, пробковидного, шаровидного и зубчатого ядра.
Ядро шатра получает информацию от медиальной зоны коры мозжечка и связано с ядром Дейтерса и РФ продолговатого и среднего мозга. Отсюда сигналы идут по ретикулоспинальному пути к мотонейронам спинного мозга.
Промежуточная кора мозжечка проецируется на пробковидное и шаровидное ядра. От них связи идут в средний мозг к красному ядру, далее в спинной мозг по руброспинальному пути. Второй путь от промежуточного ядра идет к таламусу и далее в двигательную зону коры большого мозга.
Зубчатое ядро, получая информацию от латеральной зоны коры мозжечка, связано с таламусом, а через него — с моторной зоной коры большого мозга.
Мозжечковый контроль двигательной активности. Эфферентные сигналы из мозжечка к спинному мозгу регулируют силу мышечных сокращений, обеспечивают способность к длительному тоническому сокращению мышц, способность сохранять оптимальный тонус мышц в покое или при движениях, соразмерять произвольные движения с целью этого движения, быстро переходить от сгибания к разгибанию и наоборот.
Мозжечок обеспечивает синергию сокращений разных мышц при сложных движениях. Например, делая шаг при ходьбе, человек заносит вперед ногу, одновременно центр тяжести туловища переносится вперед при участии мышц спины. В тех случаях, когда мозжечок не выполняет своей регуляторной функции, у человека наблюдаются расстройства двигательных функций, что выражается следующими симптомами.
1) астения (astenia — слабость) — снижение силы мышечного сокращения, быстрая утомляемость мышц;
2) астазия (astasia, от греч.а — не, stasia — стояние) — утрата способности к длительному сокращению мышц, что затрудняет стояние, сидение и т. д.;
3) дистония (distonia — нарушение тонуса) — непроизвольное повышение или понижение тонуса мышц;
4) тремор (tremor — дрожание) — дрожание пальцев рук, кистей, головы в покое; этот тремор усиливается при движении;
5) дисметрия (dismetria — нарушение меры) — расстройство равномерности движений, выражающееся либо в излишнем, либо недостаточном движении. Больной пытается взять предмет со стола и проносит руку за предмет (гиперметрия) или не доносит ее до предмета (гипометрия);
6) атаксия (ataksia, от греч.а — отрицание, taksia — порядок) — нарушение координации движений. Здесь ярче всего проявляется невозможность выполнения движений в нужном порядке, в определенной последовательности. Проявлениями атаксии являются так же адиадохокинез, асинергия, пьяная-шаткая походка. При адиадохокинезе человек не способен быстро вращать ладони вниз—вверх. При асинергии мышц он не способен сесть из положения лежа без помощи рук. Пьяная походка характеризуется тем, что человек ходит, широко расставив ноги, шатаясь из стороны в сторону от линии ходьбы. Врожденных двигательных актов у человека не так уж много (например, сосание), большинство же движений он выучивает в течение жизни и они становятся авто

