 2014-02-03
2014-02-03 4365
4365Конъюгация — перенос генетического материала от одной бактериальной клетки (донора) к другой (реципиенту) при их непосредственном контакте. Процесс конъюгации у бактерий обнаружили Дж. Ледерберг и Э. Татум в 1946 г. Они провели следующий эксперимент. Были отобраны два ауксотрофных мутантных штамма Е. coli К-12: не способный синтезировать метионин и биотин штамм Met"" Bio"~ и не способный синтезировать треонин и лейцин штамм Thr~ Leu"~. Оба штамма в течение ночи выращивали вместе на полноценной среде. Затем смешанную культуру центрифугировали, отмывали от полноценной среды и высевали на минимальную питательную среду. На минимальной питательной среде без метионина, биотина, треонина и лейцина появились прототрофные колонии Met+ Bio+ Thr+ Leu+ с частотой около 1 на каждые 107 клеток. Дополнительные опыты показали, что ни трансформации, ни трансдукции в данном случае не происходило. Из этого следовало, что образование рекомбинант-ных геномов происходило в результате контакта родительских клеток. Вскоре были получены микрофотографии конъюгирую-щих бактерий кишечной палочки, которые свидетельствовали о том, что между бактериями при конъюгации образуется цито-плазматический мостик.
В 1952 г. Хейс установил неравноценную роль родительских штаммов при конъюгации. Выяснилось, что один штамм является донором (мужским), другой — реципиентом (женским). Клетки-доноры обладают половым фактором F. Он является конъюгатив-ной плазмидой и представляет собой циркулярно замкнутую молекулу ДНК. Половой фактор F автономно существует в цитоплазме. Бактериальные клетки с фактором F обозначают F4", а не имеющие его — F^~. В составе генома конъюгативной плазмиды имеется tra-оперон, гены которого контролируют образование половых ворсинок (пилей) донорской клетки, необходимых для осуществления контакта с реципиентной клеткой, конъюга-тивнйй перенос собственной плазмиды или хромосомной ДНК, а также репликацию автономной плазмиды.
Механизм переноса генетического материала при конъюгации из бактерии донора в бактерию реципиента показали В. Вольман и Ф. Жакоб. При конъюгации фактор F может перейти из мужской в женскую клетку и превратить ее в F4". Доноры F4" переносят довольно эффективно F-плазмиду во все клетки F~~, a гены хромосомы — с низкой частотой (10"~5).
Половой фактор F обладает способностью включаться в геном бактерии и тогда из цитоплазматической структуры превращается в фрагмент хромосомы. Клетки, в которых возникает этот процесс, образуют Hfr-штамм. Доноры Hfr переносят бактериальную хромосому с фиксированной точки — сайта интеграции плазмиды, ориентированным образом и с высокой частотой (10—2— 10 3). Интегрированный F-фактор переносится последним. Генетическим методом идентифицировано около 25—30 сайтов интеграции фактора F в хромосому. При конъюгации клетки-доноры F1" или Hfr соединяются с клетками-реципиентами F~ при помощи конъюгационного мостика — особой протоплазматической трубки, образуемой клеткой F4". В клетке донора Hfr под влиянием фермента эндонуклеазы в точке внедрения фактора F происходит разрыв цепи ДНК. Свободный конец одной из цепей ДНК постепенно начинает передвигаться через конъюгационный мостик в клетку реципиента (F"~) и сразу же достраиваться до двух-цепочной структуры. На оставшейся в клетке-доноре цепи ДНК синтезируется вторая цепь.
Так как фактор F у разных штаммов Hfr включается в хромосому и разрывает ее в разных местах, переход хромосом в реци-пиентную клетку начинается с разных участков. Для переноса всей цепи ДНК в клетку реципиента требуется при 37 "С
100 мин, но конъюгационный мостик очень хрупкий, легко разрывается, и, как правило, вся цепь не успевает перейти. Поэтому с более высокой частотой передаются гены, расположенные около начальной 0-й точки хромосомы донора. Затем ДНК донора в гомологичных участках вступает в контакт с ДНК реципиента, и в результате кроссинговера некоторые участки одной цепи ДНК реципиента заменяются фрагментами ДНК донора.
Искусственное прерывание конъюгации через определенное время после начала скрещивания и выявление рекомбинантов дали возможность определить последовательность перехода разных генов донора в клетку F~. На основании определения времени передвижения фрагментов разной длины из клеток Hfr в клетки F~ было установлено расстояние между генами в минутах, что позволило построить карты хромосом.
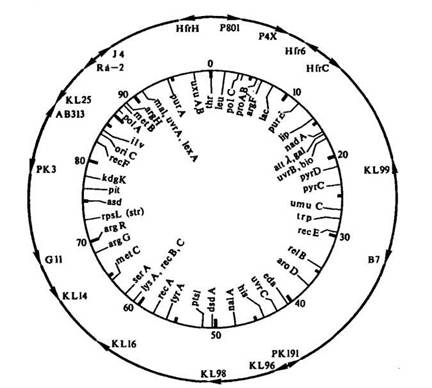
Рве 23. Неполная кольцевая карта хромосомы Е. соН К-12
В основе построения карт хромосом лежат последовательность расположения генов в хромосоме и расстояние между ними в минутах. Вся окружность хромосомы Е. coli составляет 100 мин. К настоящему времени на карту Е. coli К-12 нанесено более 1000 генов, что составляет около 30 % ее генетической емкости (рис. 23). Иногда включенный в хромосому Hfr половой фактор освобождается и при этом (подобно профагу) может захватить с собой прилегающий к нему участок ДНК бактерии. При конъюгации половой фактор вместе с.фрагментом ДНК иногда переходит в женскую клетку, превращая ее в мужскую и передавая ей свойства, контролируемые фрагментом хромосомы донора. Процесс переноса генетической информации при помощи полового фактора называется сексдукцией.








