 2015-07-14
2015-07-14 2089
2089(Н. А. Тушмалова, 1987, с дополнениями)
| Группы животных | Ароморфозы | |
| морфологический уровень (организация нервной системы) | функциональный уровень (высшая форма физиологических механизмов поведения) | |
| Простейшие | Донервный уровень | Привыкание,сенситизация |
| Кишечнополостные | Примитивная диффузная нервная система | Привыкание + тренированность |
| Плоские черви | Зачатки цефализации | Нестойкие условные рефлексы |
| Кольчатые черви | Ассоциативный центр мозга — грибовидные тела | Истинные условные рефлексы |
| Насекомые | Структурно-функциональная дифференцировка головного мозга | Положительные и отрицательные условные рефлексы, инструментальные условные рефлексы, цепные и комплексные условные рефлексы, условные рефлексы на отношения |
ПОЗВОНОЧНЫЕ
Прежде чем перейти к анализу путей развития форм обучения у позвоночных, вкратце остановимся на рассмотрении особенностей поведения у самых низших хордовых — бесчерепных и круглоротых.
Бесчерепные. Б. Ф. Сергеевым (1967) показано, что у представителя бесчерепных — ланцетника центральная нервная система построена по типу нервной трубки, характеризуется эквипотенциалъностъю, то есть отсутствием выраженной дифференциации и цефализации, причем специализированные рецепторы еще не развиты. Если производить сочетание слабого условного раздражителя (света) с тем же светом, но большей интенсивности или на слабое тактильное раздражение, подкрепляемое электрическим ударом, то у этих животных удается выработать активную избегательную реакцию. Она сохраняется в течение 10 ч, угасая уже на вторые сутки, ее каждый день приходится вырабатывать заново. Создается впечатление, что у бесчерепных намечаются признаки ассоциативного условнорефлекторного обучения, однако оно протекает по типу повышения возбудимости к условному раздражителю, то есть сходно с суммацион-ными рефлексами.
У типичного представителя круглоротых — речных миног впервые в эволюции формируются основные рецеп-торные системы и в зачаточном состоянии закладываются все подразделения мозга. Головной мозг состоит из конечного, промежуточного, среднего и ромбовидного вместе с мозжечком. В результате исследований А. В. Бару, В. В. Фанарджяна и Б. Ф. Сергеева было выявлено, что у миног вырабатываются оборонительные условные рефлексы после 3—4 сочетаний разномодаль-ных сигналов (свет + электрический ток), эти реакции сохраняются в течение лишь одного дня, восстановление происходит самостоятельно или после нескольких подкреплений. Иными словами, хотя у круглоротых ассоциативное обучение носит характер условнорефлектор-ной связи, этот тип реакций по ряду признаков напоминает еще суммационные рефлексы.
У пластинчатожаберных рыб дифференциация структур переднего мозга в морфофункциональном отношении еще недостаточно развита (А. И. Карамян, И. В. Ма-люкова, 1987). Это отражается и на характере условно-рефлекторной деятельности. У акул и скатов условные рефлексы вырабатываются медленно, осуществляются с большим латентным периодом, характеризуются непрочностью, значительной вариабельностью выполнения. Функцию высшего интегративного мозгового аппарата у этих животных выполняет сильно развитый мозжечок с его богатыми межцентральными связями с остальными отделами мозга. Передний мозг в условнорефлекторной деятельности участия еще не принимает.
Костистые рыбы отличаются более сложной и дифференцированной организацией головного мозга и сенсорных систем. V них вырабатываются истинные пищевые и оборонительные условные рефлексы на простые и сложные световые, звуковые, обонятельные сигналы, постоянное и переменное магнитное поле. Костистые рыбы характеризуются всем репертуаром форм ассоциативного научения и сенсорного различения. Однако, как подчеркивает Б. Ф. Сергеев (1967), разобщенность центральных аппаратов сенсорных систем, а также слабость нервных процессов препятствует образованию внутри- и межсенсорных условных рефлексов (ассоциативные вре-
147
менные связи), что свидетельствует о низком уровне ана-литико-синтетической деятельности.
В эволюции позвоночных рептилии занимают особое положение, ибо они представляют собой тот узловой этап эволюции, от которого формируются две независимые филогенетические линии развития мозга: стриар-ная, представленная птицами, и кортикальная, к которой относятся все млекопитающие вплоть до приматов и человека.
Мозг рептилий характеризуется прогрессивным усложнением структурной организации, и впервые возникает примитивная таламокортикальная система связей (М. Г. Белехова, 1977). Обнаружена слабая способность к фиксации следов выработанных условных рефлексов: при обучении новой задаче стирались следы предыдущего обучения. Стриатуму у черепах приписывается более важная роль в интегративной деятельности мозга, чем дорсальной коре, имеющей еще достаточно примитивную организацию.
МЛЕКОПИТАЮЩИЕ
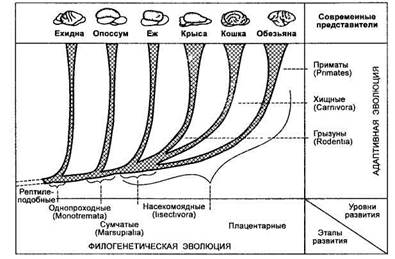
Обратимся к филогенетическому древу (рис. 18), из которого видно, что по мере развития из рептилеподоб-ных предков основного ствола млекопитающих от него отходит ветвь современных однопроходных, позже сумчатых. Стволом же плацентарных млекопитающих являются насекомоядные, от которых произошли приматы, хищные и грызуны. Большая часть отрядов современных млекопитающих — результат параллельной эволюции, а не последовательной, к которой можно было бы отнести однопроходных, сумчатых, насекомоядных. В этом ряду и далее — вплоть до высших приматов и гоминид могут быть прослежены эволюционные преобразования мозга и поведения.
Филогенетический подход не исключает и другого, чрезвычайно важного адаптационного подхода, который основан на изучении ныне живущих форм, обладающих разной степенью адаптации (специализации) в смысле развития мозга и сенсорных систем. Такие животные не обязательно должны иметь общую линию эволюции, и, следовательно, выводы, сделанные в результате такого
Рис. 18
Эволюционное древо млекопитающих с выделением этапов
филогенетической эволюции и уровней адаптивной эволюции
(по А. С. Батуеву, 1979)
сравнения, будут относиться только к общим принципам адаптации и выживания. Эти исследования углубляют понимание связи между развитием структурных образований мозга и их участием в целостном поведении. Филогенетический и адаптационный подходы взаимно дополняют друг друга. Поэтому правомерно говорить о филогенетической и адаптивной эволюции. Понятно, что сравнение особенностей поведения и конструкции мозга у грызунов, хищных, приматов не имеет никакого отношения к филогенетической эволюции, а лишь к адаптивной в смысле усложнения организации мозга и усовершенствования биологических адаптации. Начиная от насекомоядных можно проследить начальные этапы формирования всех таламических ядерных образований и проекционных зон неокортекса, характерных для высших млекопитающих. У насекомоядных (ежей) впервые в филогенезе формируется ассоциативная таламокортикалъная система, которой присущи черты примитивной организации (А. С. Батуев, 1981) (см. гл. 9). Б. Ф. Сергееву (1967) удалось выработать у
ежей ассоциативные временные связи возбудительного и тормозного характера путем сочетания разномодальных звуковых и зрительных раздражений. И. В. Малюкова (1981) установила у ежей с помощью сложных двигательных условных рефлексов различение освещенности, полос разной ориентации, фигур. Такие условные рефлексы вырабатывались быстрее, чем у черепах, и сохранялись в течение 40-50 дней. Научившись новой задаче, ежи не забывали предыдущих навыков, что свидетельствует о большой прочности хранения следов.
Однако условные рефлексы на одновременный и последовательный комплексные раздражители, которые вырабатывались с трудом, часто сопровождались невротическими состояниями. В отличие от высших млекопитающих у ежей слабо развиты ориентировочно-исследовательские реакции, не наблюдается использования метода «проб и ошибок», их нервные процессы характеризуются инертностью. Наряду с участием примитивной ассоциативной коры в процессах сенсомоторного синтеза основная интегративная роль в общемозговом контроле за сложными формами поведения принадлежит подкорковым отделам переднего мозга — стриарным образованиям. По этому признаку насекомоядные еще сохраняют черты, присущие рептилиям.
ПРИМАТЫ
Важнейшим показателем эволюционного прогресса нервной системы является развитие неокортекса, а в нем — ассоциативных кортикальных полей и соответствующих ядер таламуса (А. С. Батуев, 1981). Именно эти структуры мозга достигают у обезьян высокого уровня развития. В самой коре впервые в эволюции формируются длинные пучки волокон, связывающие различные области коры между собой. Все это создает предпосылки для выполнения весьма сложных форм поведения.
Значительное место в жизни обезьян имеет ориентировочно-исследовательская деятельность, которая, по мысли Я. Дембовского (1965), является зачатком будущего интеллекта. Убедительно показано, что обезьяны способны к формированию самых сложных форм услов-норефлекторной деятельности, связанной с тонким
различением зрительных объектов по самым разнообразным признакам. Исследования следовых условных реф-. лексов выявили прогресс образной памяти. Длительность хранения следов у низших обезьян исчисляется месяцами и годами. Обезьянам присущи все формы когнитивного обучения: образное поведение, рассудочная деятельность и вероятностное прогнозирование.
Все вышесказанное позволило А. И. Карамяну (1976), А. И. Карамяну и И. В. Малюковой (1987) выдвинуть представление о существовании критических этапов развития мозгового обеспечения поведения в эволюции позвоночных.
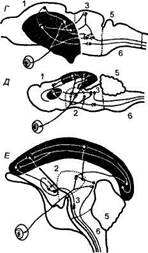 Первый критический этап представлен бесчерепными и круглоротыми, характеризующимися еще слабой дифференциацией головного мозга (рис. 19). Так, у лан-
Первый критический этап представлен бесчерепными и круглоротыми, характеризующимися еще слабой дифференциацией головного мозга (рис. 19). Так, у лан-
Рис. 19
Этапы развития уровней интеграции в филогенезе позвоночных (по А. И. Карамяну, 1976)
а — бесчерепные (ланцетник), епинальный уровень интеграции, в центре гигантская клетка с аксоном, обеспечивающим нисходящие связи; б — круглорогые (минога), бульбомезэнцефалический уровень интеграции; в — пластиножаберные рыбы (акула), мезэнцефалоцеребеллярный уровень интеграции; г — рептилии (черепаха), ди-энцефалотелэнцефальный уровень интеграции; д — насекомоядные (еж), стриокор-тикальный уровень интеграции; е — приматы (обезьяна), неокортикальный уровень интеграции; заштрихованы высшие интегративные уровни мозга; 1 — конечный мозг; 2 — промежуточный; 3 — средний; 4 -- задний; 5 — мозжечок; 6 — продолговатый мозг. На схеме показаны только зрительные афферентные проекции. Относительные размеры мозга разных животных не соблюдены.
 цетников (бесчерепные) удается выработать лишь сум-мационные реакции, а у миног (круглоротые) — квазиусловные рефлексы, занимающие промежуточное положение между суммационными и истинными рефлексами.
цетников (бесчерепные) удается выработать лишь сум-мационные реакции, а у миног (круглоротые) — квазиусловные рефлексы, занимающие промежуточное положение между суммационными и истинными рефлексами.
Второй критический этап представлен пластиножа-берными рыбами (скаты, акулы), структуры таламуса и конечного мозга которых еще недостаточно развиты. У этих животных удалось выработать условные рефлексы, которые были непрочными, сохранялись в течение нескольких суток и характеризовались невысоким уровнем их осуществления. Передний мозг пластиножабер-ных рыб не принимает участия в организации сложных поведенческих актов.
Третий этап представлен рептилиями (особенно черепахами), на поверхности конечного мозга которых формируется примитивный неокортекс. Эти животные способны к формированию более сложных форм поведения по сравнению с низшими позвоночными. Однако высшая нервная деятельность черепах отличается слабостью нервных процессов, несовершенством памяти, отсутствием способности к вероятностному прогнозированию. Ведущую роль в интегративной функции мозга у черепах выполняет полосатое тело (стриатум).
Четвертый критический этап представлен низшими, млекопитающими — насекомоядными. Мозг ежей характеризуется увеличением общей поверхности неокортекса и развитием дорзальных ядер таламуса. Однако четкая дифференциация структур таламуса и коры выражена еще слабо, хотя обнаружено существование примитивной ассоциативной системы.
 У ежей образуются сложные последовательные цепи двигательных поведенческих актов при зрительном распознавании. Впервые в эволюции позвоночных проявляется, хотя и в слабой форме, свойство вероятностного прогнозирования. Высшая нервная деятельность ежей характеризуется примитивностью: инертностью нервных процессов, трудностью формирования межсенсорных синтезов, слабостью ориентировочно-исследовательских реакций.
У ежей образуются сложные последовательные цепи двигательных поведенческих актов при зрительном распознавании. Впервые в эволюции позвоночных проявляется, хотя и в слабой форме, свойство вероятностного прогнозирования. Высшая нервная деятельность ежей характеризуется примитивностью: инертностью нервных процессов, трудностью формирования межсенсорных синтезов, слабостью ориентировочно-исследовательских реакций.
Пятый критический этап представлен приматами, где главной особенностью организации мозга является
мощное развитие его ассоциативных систем, объединенных кортикокор-тикальными путями в единую интег-ративную систему мозга. Таким образом, создаются все предпосылки для формирования более сложных форм поведения, которые оказываются специфически присущими лишь приматам.
Высшая нервная деятельность человекообразных обезьян. Некоторые исследователи поведения животных считают, что высшие психические функции не что иное, как примитивное мышление, которым животные могут пользоваться без предварительного научения, то есть не используя прежний жизненный опыт. Так, В. Келлер после наблюдения за поведением шимпанзе полагал, что высшие психические функции являются выражением изначальных свойств и способностей мозга.
И. П. Павлов и его сотрудники, повторив опыты Келлера, пришли к иному выводу. Шимпанзе составляет пирамиду из ящиков, чтобы достать гроздь бананов, подвешенных к потолку, на основании перенесения в новую ситуацию старого опыта, на основании проб и ошибок (рис. 20). Сам И. П. Павлов писал по этому поводу: «...Каждая новая ассоциация, касающаяся отношения внешних вещей, есть прибавление знания, а пользование этим знанием есть то, что называется пониманием».1
Еще Ч. Дарвин (1871) отмечал, что подражание сыграло значительную роль в процессе антропогенеза, при-
1 Неопубликованные и малоизвестные материалы И. П. Павлова. Л., 1975. С. 96.
чем у родоначальников человека подражание в сочетании с рассудком сыграли исключительную роль в выборе оптимальных решений.
Л. А. Фирсов (1987) выделяет подражательную деятельность в качестве самостоятельной формы поведения антропоидов, которая оказывается особенно выраженной у детенышей шимпанзе в возрасте 2—2,5 лет. Подражание после однократного наблюдения решения проблемных задач или демонстрации дифференцирования предъявляемых сигналов особью-«актером» сохраняется у «зрителя» от нескольких часов до нескольких недель. Причем кроме скорости и прочности подражания отмечается его очень широкий диапазон.
Механизм подражания следует рассматривать как особую форму поведения или отражения действительности, где принцип «животное — окружающий мир» заменяется принципом «животное — другое животное — окружающий мир». Причем особь-«актер» воспринимается особью-«зрителем» как обобщенная система натуральных раздражений, которыми и являются акты поведения.
Предметная и орудийная деятельность антропоидов — это одна из базовых проблем антропогенеза. Установлено, что манипулирование разнообразными предметами среды — предметная деятельность — это неотъемлемая черта сложного поведения всех представителей отряда приматов. Антропоиды обладают способностью к целенаправленному манипулированию с предметами. В процессе этой деятельности они не только используют предмет как таковой или при необходимости изменяют его, но способны также сочленить несколько предметов в устойчивую конструкцию. Использование предметов для игры молодыми шимпанзе начинается с 2,5-3-месячного возраста. Но только в возрасте около 2,5 лет детеныши шимпанзе научались составлять новые конструкции и использовать их для приближения приманки. То же наблюдается и у других антропоидов (рис. 21).
Орудийная деятельность отчетливо проявляется у шимпанзе в опытах с огнем, который служил препятствием для достижения приманки. В опытах И. П. Павлова
 154
154
Рафаэль взял бутылку с водой и вылил ее на пламя, а в опытах Л. А. Фирсова Каролина накрыла огонь половой тряпкой и легко завладела апельсином. Из этих опытов ясно видно свойство обобщения сигнальной ценности окружающих предметов. Наблюдения за поведением шимпанзе, проведенные Л. А. Фирсовым, дают основание считать, что, перед началом каких-либо действий, у шимпанзе имеется ближайший план, ради которого они реализуются. Вот один из примеров. Подростки шимпанзе Лада и Нева, воспользовавшись ошибкой лаборантки, достали связку ключей, забытую ею на столе в 3 м от вольера, и оказались на свободе. Действия обезьян при этом были следующими: 1) отгрызание края столешницы от стола, который уже давно находился в вольере; 2) с помощью получившейся палки подтягивание занавеси и овладение ею; 3) набрасывание как лассо занавеси на стол с ключами и очень осторожное подтягивание связки ключей к вольере; 4) открывание висячего замка и выход на свободу.
Этот пример отражает реализацию уже принятого окончательного решения, которое сформировалось как на основе текущей деятельности, так и имеющегося у обезьян жизненного опыта. В любом случае обезьяна способна в известной степени предвидеть результаты своих действий.
Орудийная деятельность основана на анализе связей между предметами, явлениями и результатами манипулирования с предметами. При этом важную роль играет уровень обобщения элементов среды с тем, чтобы можно было применить любой предмет в любой ситуации, придерживаясь логики принятого решения. Целенаправленность поведения с обобщенным использованием предметов внешней среды, с частичным их изменением
и составлением из них устойчивых конструкции является наиболее поздним эволюционным приобретением. Она обязательно проявляется в поведении ребенка, и на ней в значительной мере основывается трудовая деятельность человека. Новая ступень предметной деятельности — орудийная деятельность — развивается в процессе все большей социализации древнейшего человека.
Учитывая семейно-групповой образ жизни антропоидов, исследователи уделяют значительное внимание способам общения животных между собой и с экспериментатором. Основное внимание уделяется голосовым и жестикуляторным проявлениям. Л. А. Фирсов и В. В. Плотников (1981) отмечают выраженную «полисе-мантичность» голосовых реакций обезьян и большое значение окружающей ситуации для придания им определенной сигнальной значимости. В силу высокоразвитых свойств подражания антропоиды легко усваивают новые звуковые проявления, обогащая их экологическое значение. В определенных условиях эти артифициальные сигналы могли оказаться более точными, чем врожденные, и соответственно их биологическая роль возрастала.
Своеобразная форма общения шимпанзе с экспериментатором была изучена А. И. Счастным (1972). Обезьян приучали возвращать экспериментатору выданные им во время опыта жетоны: за возвращение круглого жетона давали обезьяне конфету, квадратного — орех, прямоугольного — компот, шестиугольного — игрушку, за треугольник — ничего не давали. Обезьяны, используя условные «знаки», сигнализировали экспериментатору и друг другу о своей потребности в пище, питье или игрушке.
Та же цель преследовалась опытом супругов Гарднеров, Премаков и других исследователей на детенышах шимпанзе и горилл. В качестве символов могли использоваться либо специальные фишки, либо азбука глухонемых. Важно то, что выученными жестами или фишками шимпанзе могли пользоваться между собой, а новички по механизму подражания 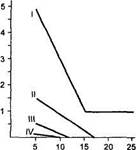 воспринимали их у обученных и успешно ими пользовались.
воспринимали их у обученных и успешно ими пользовались.
Антропоиды обладают способностью к переносу первичного научения для формирования обобщения с отвлечением от конкретных признаков сигнала. Стало
очевидным, что шимпанзе способны абстрагироваться от некоторых существенных признаков предметов или явлений и обнаруживают выраженную способность к ассоциативному мышлению. Обобщение и абстракция являются результатом чувственного опыта животного, содержат информацию об истории процесса, оценке ситуации, решении, его реализации, результатах комбинаторики старых навыков для нового решения. Поэтому обобщение — функциональный блок систематизированной информации, хранящейся в аппаратах памяти.
Вероятно, на основе сложных обобщений у древних приматов формировался знаковый язык. Л. А. Фирсов (1982) понимает под термином «язык» все формы адаптивного поведения животных и человека, в том числе врожденные коммуникативные сигналы. Это первичный язык. Знаковая система общения у обезьян позволяет различать во вторичных языках стадию «А» (довербальные понятия) и стадию «Б» (вербальные понятия). Последние, как правило, реализуются с помощью речевого аппарата. Соотношение классификации языков приводится в табл. 4.
Существенной стороной поведения антропоидов являются формы их сосуществования в сообществах (семьях, группах, стадах). Такую кооперацию можно рассматривать как присущий приматам тип зоосоциального поведения, при котором действия нескольких особей объединяются для выполнения какой-либо операции, направленной на достижение полезного результата. Ко-оперативность поведения обезьян зависит прежде всего от иерархических рангов партнеров, а затем и от других факторов, таких как уровень мотивации, физиологическое состояние особей и др.
Кроме обычного наблюдения за поведением животных Л. А. Фирсовым был использован весьма показатель-таблица 4 Классификация языков (уровни отражения) (Л. А. Фирсов, 1987)
| И П Павлов | Л А Орбелн (1949) | Л А Фирсов (1974) | |
| 1 сигнальная система | I сигнальная система | Допонятииный уровень | Первичный язык |
| Промежуточный этап | Стадия «А» довербальные понятия | Вторичный язык | |
| 11 сишапьнэя система | I! сигнальная система | Стад/я «Б» вербальные понятия | Тоже |
ный объективный тест на кооперативное поведение двух обезьян. Возле одной из кормушек находился рычаг, нажатие на который открывало доступ к пище в противоположной кормушке, и наоборот. Кооперативное поведение устанавливается тогда, когда обезьяны по очереди подкрепляют одна другую. Однако такое поведение разрушалось, как только проявилось доминирование одной особи и подчиненность другой. В этом случае подчиненная особь в несколько раз чаще нажимала на рычаг, чтобы накормить своего партнера, чем доминирующая.
Наконец, хорошо изучена способность антропоидов к длительному хранению следов. Главной отличительной особенностью шимпанзе является широкое использование образной памяти (рис. 22).
Вышеприведенные данные об уровне психического развития антропоидов, включающего подражательную, орудийную, ориентировочно-исследовательскую деятельность, способность к обобщению и абстрагированию сигналов, использованию знаковой системы сигнализации и высокого уровня образной памяти, позволяют заключить, что обезьянообразные, ближайшие предшественники древнейшего человека, обладали не меньшим развитием перечисленных функций.
§ 21. ФОРМИРОВАНИЕ ПОВЕДЕНИЯ В ОНТОГЕНЕЗЕ
Типы онтогенеза. Переход от автоматизированных реакций к целенаправленным происходит постепенно и определяется зрелостью центральной нервной системы. Упреждающий характер развития поведения заключается в готовности ряда систем организма к определенной деятельности еще до перехода к самостоятельной жизни.
Рис 22
Состояние
образной памяти
(отсроченный выбор)
на фоне тренировки
различения
зрительных сигналов
(по Л. А. Фирсову,
1987)
По оси ординат — отсрочка в критическом испытании, мин, по оси абсцисс — номера кри тического испытания после образования условного рефлек са, / — шимпанзе, // — пави аны, III — макаки, IV - ка пуцины
В. Г. Кассиль (1987) отмечает, что пищевое поведение на ранних этапах жизни регулируется иначе, чем в более поздние периоды развития. Например, у новорожденных крыс и собак отсутствует связь между активностью сосания и поступлением в организм материнского молока. При этом сигналы от рецепторов желудка менее значимы, чем у старших особей. Потребление пищи зависит в большей степени от сенсорных факторов, чем от метаболических. Питьевое поведение формируется позже, чем пищевое.
Даже у незрелорождающихся животных в самые первые этапы постнатальнои жизни большое значение имеет обучение. Например, целенаправленная пищевая реакция на обонятельный сигнал формируется по принципу импринтинга, то есть запечатлевания. И только позже в связи с совершенствованием механизмов памяти начинают вырабатываться типичные условные рефлексы.
Существование только двух источников, определяющих видоспецифические особенности поведения организма — генотипа и среды, не дает достаточных оснований для разделения на две противоположные и взаимоисключающие формы сложного адаптивного поведения. Жесткая дифференциация только двух факторов, влияющих на развитие поведения — генетических и средовых, столь же трудна, как решение вопроса, от чего больше зависит площадь — от длины или от ширины (Д. О. Хебб, 1949). Еще более неплодотворна идея деления поведения на врожденное и приобретенное.
К моменту первого своего проявления любая функциональная организация является незрелой, базирующейся на врожденных реакциях. Одни формы поведения, например пищевое, высоко организовано уже к моменту рождения, другие (половое, материнское) впервые проявляются на более поздних стадиях онтогенеза.
Л. А. Орбели (1961) следующим образом охарактеризовал два типа онтогенеза животных — зрелорождаю-щихся и незрелорождающихся: «Одни животные уже внутри яйца, во внутриутробной жизни достигают довольно большой зрелости и попадают под влияние внешней среды, родившись или вылупившись из яйца уже вполне сформировавшимися, и им остается только при-
обретать новые условные рефлексы, а другие рождаются на свет еще не созревшими, еще не все безусловные рефлексы у них возникли и развились, а уже на них действует внешняя среда и устанавливает временные связи с теми реакциями, которые имеются уже налицо или которые вот-вот только развиваются, и в зависимости от этого вся безусловнорефлекторная деятельность уже дальше протекает иначе, чем она протекала бы, если бы животное было ограждено от влияния внешней среды. Этот „переплет", который создается между врожденными рефлексами и наслаивающимися на них условными рефлексами, „переплет" не просто суммирования, накладывания друг на друга, а проникновения друг в друга и взаимного влияния, представляет чрезвычайно важную сторону физиологии нервной системы и не менее важную сторону биологического процесса приспособления тех или иных видов к тем условиям существования, в которых они должны жить».1
Формирование пищевого поведения. Движения, характерные для пищевого поведения у новорожденных млекопитающих, стереотипны и в значительной степени не зависят от мотивационных факторов. В то же время специфическая пищевая реакция новорожденных (сосательные движения, слюноотделение) до первого кормления может быть вызвана тепловым воздействием. Эта реакция усиливается при прикосновении к мордочке пучком щетинок, имитирующим шерстный покров. В опытах X. Харлоу и Р. Циммермана (1959) детенышей макак-резусов изолировали от матерей и кормление их производили с использованием модели матери в форме проволочного цилиндра или такого же цилиндра, но обтянутого мягкой тканью. Независимо от того, какая модель применялась для кормления, детеныши большую часть времени проводили в контакте с «матерчатой матерью». Отсутствие стабильной температуры тела детеныша в ранний период развития компенсируется теплом матери.
Большинство исследователей считают, что для незре-лорождающихся детенышей млекопитающих наиболее
1 Орбели Л. А. Собр. соч. 1964. Т. 1. С. 382.
общим положительным раздражителем, удерживающим новорожденных около матери и обеспечивающим первый контакт с нею путем пищевых реакций, является теплая поверхность. Иными словами, наиболее рано проявляющейся в онтогенезе врожденной реакцией детенышей млекопитающих является положительный термотаксис, который и обеспечивает контакт с матерью в первые минуты жизни.
У зрелорождающихся детенышей млекопитающих (например, у копытных и хоботных), где из-за передвижения всего стада или молодняка с самками отсутствует постоянная гнездовая территория, наряду с прикосновением к теплой поверхности, которое предшествует первому кормлению, важное место среди врожденных рефлексов занимают ответы на зрительные сигналы, такие, как затемнение над головой, имитирующее мать, стоящую над детенышем (А. Д. Слоним, 1976), и следование за движущимся предметом. Эти рефлексы хорошо проявляются на 1—2-й день жизни. Но если кормление происходит из рожка без затемнения над головой или затемнение не подкрепляется пищей, то в течение первых трех дней положительная пищевая реакция на этот сигнал исчезает.
Формирование пищевых реакций не завершается к моменту первого кормления, а продолжается в направлении постепенного усложнения. Этот процесс облигат-ного обучения (Г. Темброк, 1964) зависит от критических периодов постнатального онтогенеза. У кошек выделяют 4 таких периода: первый — от рождения до 7-9 дней, то есть до момента открывания глаз и наружных слуховых проходов. Второй длится около 20 дней и заканчивается к месячному возрасту. Оба эти периода иногда объединяют в один — ранний постнатальный. Третий период — до 9-11-недельного возраста и четвертый — завершается к 7 мес постнатального развития.
I. Животное первого возрастного периода с недоразвитыми системами зрительного и слухового восприятия ориентируются главным образом с помощью кожно-тем-пературной и обонятельной чувствительности (К. В. Шу-лейкина, 1971). По термотактильным и обонятельным раздражениям котенок способен различать признаки
«своей» и «чужой» гнездовой территории. Комплекс таких реакций обозначают как хоминговую ориентацию детенышей. Успешность достижения домашней зоны при этом крайне низка. Поэтому постоянный контакт с матерью в этих условиях имеет большое биологическое значение. Ведущую роль в поддержании такого контакта играет мать, которая находит детенышей, ориентируясь по издаваемым ими крикам, возвращает их в гнездо и кормит (Н. Н. Соколова, 1984).
И. Во второй период начинают формироваться координированные и довольно быстрые движения глазных яблок и ушных раковин в сторону источника раздражения. Локомоторный аппарат и все сенсорные системы развиваются весьма активно. Мать, а затем и котята все чаще покидают гнездо. Именно в этом периоде кошка впервые начинает издавать призывные материнские сигналы, в ответ на которые котенок к ней приближается. Лактирующая самка издает также непрерывные акустические сигналы (тихое мурлыканье), которые также служат ориентиром для котят при поиске ими гнезда.
Зрительный контроль расположения гнезда, матери и других членов семьи отчетливо проявляется у котят лишь к месячному возрасту. Хоминговая реакция при этом заменяется ориентировочно-исследовательскими и игровыми формами поведения.
Теперь уже инициатива кормления все больше начинает зависеть от детенышей. Причем мать продолжает активно реагировать на призывные крики детенышей и обеспечивает непрерывный двусторонний контакт с ними. В этом периоде у котят впервые начинает проявляться хищничество.
III. Способность к хищничеству развивается только в третьем (ювенильном, игровом) периоде жизни. Мать все чаще покидает детенышей, даже отталкивает и избегает их. Периоды кормления становятся менее продолжительными. Интенсивно развивается ориентировочно-исследовательское поведение. Игра способствует освоению окружающей территории, отрабатывает и тренирует элементы будущих взрослых форм поведения, способствуя накоплению необходимого опыта в сенсорном и моторном планах. Причем это свойство присуще
6 Зак К 64
только высшим млекопитающим. У них в игровом периоде заметно совершенствуется способность фиксации внимания, появляется реакция слежения, наблюдения за действием матери и сопометников, иногда подражание им. При этом возникают первые попытки самостоятельного преследования добычи и ее умерщвления.
IV. Четвертый период — полового созревания у кошек заканчивается в 7—9 мес постнатального онтогенеза. Доля игровых компонентов в поведении уменьшается. Хищническое поведение совершенствуется, стереотипизирует-ся. Условнорефлекторная деятельность становится сложной и разнообразной. Непосредственное преследование добычи сменяется более сложными формами поведения, включающими элементы вероятностного прогнозирования и экстраполяции. К концу этого периода молодые котята могут самостоятельно охотиться и защитить себя, и семья распадается.
Взаимоотношение детенышей и родителей. Материнская забота о потомстве может быть определена как многоактное поведение, в котором интегрированы врожденные и индивидуально приобретенные компоненты. Формы заботы о потомстве у разных отрядов и семейств млекопитающих различны. Наибольшей выраженности среди незрелорождающихся млекопитающих забота о потомстве достигает у обезьян. Детеныш сразу после появления на свет повисает на шее матери, и она практически не расстается с ним 15-20 дней. Такому тесному контакту способствует положительный термотаксис детенышей и свойственные им хватательный и сосательный рефлексы. Начиная с недельного возраста хватательный рефлекс ослабевает, активизируется исследовательская деятельность. Большую роль в организации материнского поведения, особенно у человекообразных обезьян, играют голосовые реакции детенышей, одни из которых свидетельствуют о его состоянии, другие — об угрозе нападения, что заставляет мать немедленно мобилизоваться для защиты детеныша.
У большинства копытных, живущих стадами, детеныши пытаются встать и сделать первые шаги уже через 15-20 мин после рождения и очень скоро начинают повсюду следовать за матерью. Первые 3—4 ч являются чувст-
вительным периодом для детеныша и матери, во время которого устанавливается тесная связь между ними по принципу импринтинга. Детеныш запоминает на длительное время или на всю жизнь зрительные образы, обонятельные признаки, звуковые сигналы матери или объекта, заменившего ее. Если в течение критического периода формирования общения детеныш не имеет возможности установить связи с особями своего вида, впоследствии он может оказаться неспособным к общению с себе подобными и будет испытывать затруднения не только в коммуникативном, но и воспроизводительном поведении.
Формирование первичных коммуникативных связей устанавливается и путем «обратного запечатлевания» детенышей их родителями. При этом существенную роль играют акустические сигналы детенышей. Если в течение 3-4 ч после рождения изолировать мать от детеныша, она не сможет отличить его от других. С другой стороны, в период запечатления самки легко принимают и начинают вскармливать чужих детенышей.
М. М. Хананашвили (1978), Е. С. Петров (1988) специально подчеркивают значение фактора общения со сверстниками в период раннего онтогенеза для формирования свойств высшей нервной деятельности животного. Оказалось, что у животных, выращенных в условиях даже частичной внутривидовой изоляции, значительно страдает способность фиксировать следы в краткосрочной памяти, нарушается способность удержания в долгосрочной памяти нескольких систем условных рефлексов. Отмечено, что животные, выросшие в условиях такой частичной социальной депривации, характеризуются резкими отклонениями в протекании эмоциональных реакций: имеют место эмоциональная тупость, чрезмерная выраженность эмоции страха, неадекватность знака эмоции на различные раздражители.
Системогенез и теория диссолюции. Из многих теоретических представлений о возрастной динамике формирования целостного поведения выдержали проверку временем лишь две, которые мы здесь и рассмотрим.
Л. А. Орбели (1964) считал, что сопоставление филогенеза с онтогенезом (закон рекапитуляции) должно быть весьма продуктивным, ибо раскрывает картину
формирования рефлекторной деятельности: как возникают, наслаиваются друг на друга, переплетаются друг с другом, взаимодействуют, а иногда подавляют один другого различные рефлекторные акты.
В экспериментах на эмбрионах кролика и морской свинки было показано, что рефлекторные акты закладываются и выявляются уже в эмбриональном периоде, но достигают полного развития в разные периоды постна-тального онтогенеза. Постепенно включающиеся все новые и новые координации оказывают свое модулирующее влияние на те рефлекторные акты, которые сформировались ранее. Путь выявления этих влияний Л. А. Орбели видел в анализе клинического материала. Ряд психопатологических симптомов может быть истолкован «...как проявление известной рекапитуляции, как повторение пройденного ранее этапа, как выявление того, что в норме было замаскировано под влиянием более высоких, более молодых, более поздних уровней развития».1 Эта система взглядов и была названа теорией диссолюции. Ее справедливость подтверждается и экспериментальной физиологией: выключение более молодых мозговых образований позволяет выявить обычно маскируемые автоматизированные рефлекторные акты. Способность к имитации может быть присуща как облигатным, так и факультативным формам обучения и наиболее ярко выражена в определенный период раннего онтогенеза, а затем она не проявляется в такой степени, маскируясь множественными формами индивидуальных адаптации. Однако удаление у взрослой собаки лобных долей коры мозга превращает ее в автоматического имитатора, пассивно следующего за экспериментатором и повторяющего действия другой здоровой собаки (Н. А. Шустин, 1959).
П. К. Анохин (1968), разработав стройную систему представлений о функциональных системах, считал, что весь процесс отражения внешнего мира живыми организмами, закрепленный в филогенезе наследственными факторами, находит свое выражение в развитии зародышей человека и млекопитающих. В периоде эмбриональной жизни происходит развитие именно тех функцио-
Орбели Л. А. Собр. соч. 1964. Т. 1. С. 301-302.
нальных систем, которые необходимы для осуществления жизненно важных функций новорожденного, приспосабливающих его к внешней среде. Эта «подгонка» функциональных систем к экологии данного организма достигается гетерохронным и избирательным созреванием центральных и периферических структур, обеспечивающих животному приспособление функции к моменту его перехода в новую внешнюю среду. Такова наследственно закрепленная в филоэмбриогенезе особенность развития организма. Такому разновременному созреванию различных структур зародыша предшествует также гетерохронное и избирательное развитие мозговых структур данной функциональной системы.
Для каждого вида животного характерен свой своеобразный системогенез. Организующаяся система может функционировать прежде, чем она достигнет окончательного оформления. К этому сводится суть концепции сис-темогенеза.
Например, только что вылупившиеся птенцы грача отвечают раскрытием клюва на звук «кар-р» и на движение воздуха. Оба эти фактора непременно предшествуют приему пищи в естественных условиях жизни. Оказалось, что к моменту вылупления птенцов в их слуховом аппарате созрели лишь только те рецепторные элементы, которые способны воспринимать звук «кар-р». У других птиц-дуплянок эффективным стимулом раскрытия клюва оказалось затемнение дупла, когда мать, прилетающая с кормом, закрывает собой единственное отверстие, через которое поступает свет. Было установлено, что к моменту вылупления созрели те нейронные структуры мозга, которые обеспечивают восприятие перепада освещенности и переключают возбуждение на двигательные пути, управляющие раскрытием клюва.
Как легко можно заметить, две рассмотренные концепции онтогенеза поведения не исключают, а взаимодополняют друг друга. Причем каждая из них в большей мере отражает одну сторону процесса развития поведения. Если концепция диссолюции в большей мере аргументирована клинико-физиологическими материалами, то концепция системогенеза опирается на эмбрио-генетические закономерности.
Элементы адаптивного поведения в онтогенезе. Процессы, совершающиеся в ходе онтогенеза, во многом обнаруживают те же закономерности, что и в динамике филогенеза.
Первыми реакциями, проявляющимися на начальном этапе становления функции, являются таксисы. Таксисы считаются врожденными, генетически фиксированными реакциями, проявляющимися в ответ на ключевые раздражители, которые и определяют вектор протекания реакции. В дальнейшем они вступают в тесные сочетания с индивидуальным опытом животного.
Другим врожденным компонентом поведения являются безусловные рефлексы, обеспечивающие элементарный сенсомоторный уровень реагирования, способствующий подготовке висцеральных систем и скелетной мускулатуры к осуществлению определенных функций. Наличием таких врожденных компонентов поведения обеспечивается предварительная готовность к контакту с наиболее значимыми для выживания особи элементами внешней среды, «врожденное узнавание» этих раздражителей (Э. К. Фабри, 1976).
На начальных этапах формирования поведенческих адаптации проявляется ряд стереотипных двигательных актов — комплексы фиксированных действий. Выполнение этих действий автоматизировано, но в ходе онтогенеза происходит уточнение, дифференцировка «узнавания» тех раздражителей, которые в дальнейшем будут запускать сложные инстинктивные акты. Совершенствование взаимосвязи стимула и реакции происходит путем облигатного и факультативного обучения. Особенностью облигатного обучения является его приуроченность к определенным сенситивным (критическим) периодам онтогенеза. Эти периоды приурочены к самым начальным этапам формирования функции и, как правило, являются непродолжительными.
Облигатное обучение может осуществляться путем импринтинга и играет большую роль в становлении многих форм поведения.
Предполагается, что внутренние мозговые механизмы запечатления сходны с механизмами доминанты (А. С. Батуев, Н. Н. Соколова, 1987). Возникновение
состояния доминанты и характеризует особенности протекания чувствительного (критического) периода становления той или иной функции.
Формирование видового стереотипа поведения может происходить и путем подражания (имитации). Молодые животные путем подражания учатся выполнять некоторые жизненно необходимые действия обычного поведенческого репертуара своего вида. Имитация может осуществляться и в процессе факультативного обучения, идентифицирующего поведение в связи со случайными, преходящими факторами среды.
Путем факультативного научения в онтогенезе приобретаются сложные инструментальные условные рефлексы. При этом у молодого животного-зрителя вырабатывается определенный навык в результате одного лишь созерцания действий другой особи, направленных на решение задачи.
Одним из средств приобретения индивидуального опыта в разные периоды онтогенеза являются формы ассоциативного обучения — классические и инструментальные условные рефлексы.
В постнатальном онтогенезе описано явление периодизации развития условнорефлекторнои деятельности на основе созревания мозговых систем и метаболических процессов в организме.
Г. А. Образцова (1964) в исследованиях на собаках выделяет период от рождения до 2,5-3 недель, в котором преобладают безусловные рефлексы и рефлексы на натуральные сигналы.
В период от 3 до 6 недель появляются и упрочиваются условнорефлекторные связи со всех сенсорных систем.
В период от 6 до 12 недель жизни отмечена наибольшая скорость образования условных рефлексов, их широкая генерализация в сенсорном и моторном звеньях (см. гл. 6). В период от 6 мес и старше условнорефлек-торная деятельность приобретает выраженные индивидуальные особенности. У взрослых особей отчетливо проявляются формы образного поведения, экстраполяции и особенно вероятностного прогнозирования, что коррелирует с высоким уровнем развития интегративных систем мозга.
§ 22. ЗАКЛЮЧЕНИЕ
Поведение — один из основных факторов эволюции популяций. В основу классификации форм поведения может быть положен принцип разделения потребностей организма на биологические (витальные), социальные (зоо-социальные, ролевые) и идеальные (поведение саморазвития). Существуют биологические предпосылки сложных форм потребностей у человека. Формы индивидуального обучения подразделяются на 3 группы: 1) облигатное неассоциативное (суммация, привыкание, импринтинг, подражание); 2) факультативное ассоциативное (классические и инструментальные условные рефлексы) и 3) когнитивное (образное психонервное поведение; элементарная рассудочная деятельность; вероятностное прогнозирование).
В динамике эволюции животного мира для донервных форм организации (простейшие) индивидуальное обучение проявляется в формах привыкания и сенситизации. Появление нервной системы (кишечнополостные) связано с усложнением обучения. Возникновение ЦНС (черви, моллюски) влечет к появлению способности к ассоциативному обучению, а четкое выделение головного мозга в результате цефализации (насекомые) — сложных форм ус-ловнорефлекторного поведения. Приведенная динамика совершенствования форм обучения в основном повторяется у домлекопитающих позвоночных. У млекопитающих в связи с появлением ассоциативных систем мозга развиты все формы обучения, особо высокого совершенства достигающие у приматов (антропоиды) с их предметной и орудийной деятельностью, подражанием, образной памятью, способностью к обобщению и абстрагированию с использованием знаковых систем сигнализации в общении.
В динамике индивидуального развития особи, подчиняющегося правилам системогенеза и диссолюции, реализуется закон рекапитуляции: онтогенез поведения в сжатой форме напоминает длительный процесс эволюционного созревания поведенческих адаптации. Определяющими являются типы онтогенеза: зрелорождающие-ся и незрелорождающиеся организмы. В последнем случае созревание мозга и поведенческих проявлений может достигнуть дефинитивного уровня лишь при условии постоянного общения с родителями.
Глава 5. ФАКТОРЫ ОРГАНИЗАЦИИ ПОВЕДЕНИЯ
§ 23. ГЕНЕТИЧЕСКАЯ
ДЕТЕРМИНАЦИЯ СВОЙСТВ ПОВЕДЕНИЯ
1 енетика поведения. Генетика поведения как самостоятельная научная дисциплина существует сравнительно недавно, ее возраст исчисляется с момента выхода книги Д. Фуллера и В. Томпсона «Генетика поведения» в 1960 г. Главной стержневой задачей этой науки сразу же стало изучение генетических основ индивидуально приобретенных форм поведения.
Вся совокупность генетического материала организма или его часть, то есть комплекс всех наследственных задатков, контролирующих развитие, строение и жизнедеятельность организма, именуют генотипом. Реализация генотипа особи происходит при ее непрерывном взаимодействии с внешней средой, а продукт интеграции генотипических и средовых воздействий, заключающийся в наборе морфологических, физиологических и поведенческих признаков организма, именуется фенотипом. Между генотипом и фенотипом нет однозначного соответствия. Генотип лишь определяет возможные пути развития организма и его свойств при взаимодействии с внешней средой. Последняя влияет на фенотипическую изменчивость организма, а диапазон этой изменчивости зависит от так называемой нормы реакции, которая задается генотипом.
Напомним основные наиболее устойчивые понятия. В генах, то есть молекулах ДНК, заключены потенции организма. Эти потенции определяют норму реакции организма в отношении его физических и поведенческих функций. ДНК передает закодированное генетическое сообщение молекулам РНК. Таким образом, РНК
получает от ДНК исходные инструкции, записанные с помощью линейного кода. До появления надлежащих стимулов большая часть комплексов ДНК находится в неактивной форме. После соответствующей стимуляции ДНК активируется и быстро синтезирует РНК так, что становится возможным и синтез специфических белков. В большинстве молекул ДНК каждый участок (локус) специализирован, поэтому его изменение приводит к фенотипическому дефекту поведения в результате синтеза измененных РНК и белков.
Уже с момента возникновения оплодотворенной клетки в результате слияния двух родительских клеток начинается реализация генетической информации, полученной от этих родительских клеток. На зародыш воздействует внешняя среда, которая передает свои влияния вначале через материнский организм, а затем и непосредственно. В дальнейшем развертывание свойств генотипа в форме различных структурно-функциональных комплексов происходит под непрерывным воздействием среды. Поэтому любой фенотипический признак — это сплав из врожденных и приобретенных компонентов.
Задачи генетики поведения состоят в стремлении выделить генетическую сущность в фенотипическом проявлении поведенческих актов, попытаться направленно влиять на генотипическую сущность поведенческих проявлений и базовых свойств нервной системы и наоборот, определить силу средового воздействия на генотип организма и оценить стабильный характер возникшей поведенческой изменчивости и др.
Несмотря на свою молодость, генетика поведения не только разработала ряд специфических методов исследования, но и сформулировала ряд принципиальных теоретических положений.
Генотип и его влияние на поведение. Фактически все современные исследования показывают, что изменения в генотипе глубоко влияют на внешне наблюдаемое поведение животных и человека.
Наиболее распространен метод лабораторной селекции генетически чистых линий животных (инбредные линии), все особи которой обладают одинаковым генотипом. Если животные разных инбредных линий, выдер-
жанные в одинаковых условиях, проявляют существенные различия в поведении, то приходят к выводу о генетической обусловленности этих различий.
Так, например, путем искусственного отбора были выведены линии крыс и мышей с высокой и низкой эмоциональной активностью в тесте «открытого поля» (рис. 23). Активными называют крыс, которые, будучи
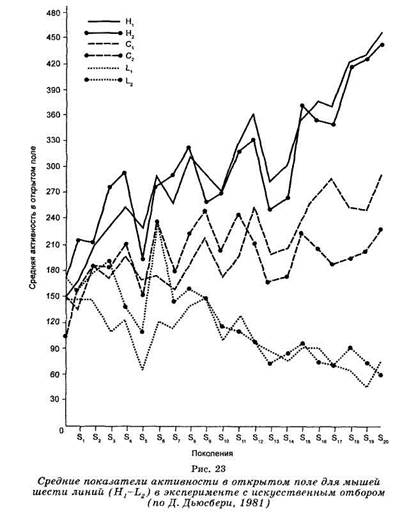
помещенными на открытую освещенную площадку, обнаруживают частые дефекации и мочеиспускания и, оставаясь неподвижными, часто распластываются на одном месте. У нереактивных крыс не было дефекации, и они свободно разгуливали по площадке. Реактивные крысы из-за высокой эмоциональности медленно обучаются избеганию удара электрического тока.
Исследования на линиях животных с известным генетическим дефектом или мутантах привели к выводу, что способность к обучению, как и любой другой признак организма, детерминирована генотипом и обусловлена действием множественных генов. Путем использования множественного скрещивания (гибридологический метод) нескольких линий мышей, различающихся способностями к обучению, А. Оливерио (1972) показал, что эти генетические различия основаны на свойствах головного мозга обеспечивать разный уровень обучения.
Р. Коллинз и Д. Фуллер (1968) вывели линию мышей, а Л. В. Крушинский и Л. Н. Молодкина (1971) — линию крыс по признаку высокой чувствительности к действию сильного звука, который вызывал у них эпилептиформные судороги. Такие генуинные формы аудиогенной эпилепсии стали широко распространенной моделью устойчивого патологического состояния. Сравнение неврологического статуса крыс линии Кру-шинского—Молодкиной с крысами линии Вистар обнаружили у первых значительный дефицит в обучении ориентировке в 12-лучевом лабиринте, процессов кратковременной памяти и общей инертности возбудительно-тормозных взаимоотношений (Е. А. Рябинская и др., 1981) (рис. 24). В то же время по скорости переделки сигнального значения условных раздражителей (положительный превращается в отрицательный и наоборот) более способными оказались крысы линии Крушинского— Молодкиной по сравнению с крысами линии Вистар (Л. В. Крушинский, 1960).
Р. Коллинз и Д. Фуллер предположили, что различия между линиями мышей по их предрасположенности к аудиогенным судорогам контролируются одним геном. Обычно один ген влияет на несколько различных признаков. Например, альбинизм контролируется одним
Рис. 24 Вероятность совершения
крысами определенного числа ошибок при поиске пищи в лучах радиального
лабиринта (по А. С. Батуеву и др., 1983) По оси ординат — вероятность; по оси абсцисс — число ошибок; 1 — средние величины у «необученных» (1-10-й опыты) и 2 — у обученных (47-56-й опыты) крыс линии Вистар; 3 — у «необученных» и 4 — у обученных крыс линии Крушинского-Молодкиной; 5 — математическая модель случайных посещений.
геном, который влияет одновременно и на признаки поведения в открытом поле. У человека гены, ответственные за микроцефалию и фенилкетонурию, вызывают расстройство в строении и биохимии мозга и сопровождаются умственной отсталостью. Однако большая часть свойств поведения животных детерминирована действием многих генов, каждый из которых в отдельности сравнительно мало влияет на фенотип.
Генетика инстинктов. Занимаясь генетикой поведения, ученые сталкиваются с необходимостью и выбором для анализа «единиц поведения». Некоторые ученые считают, что наиболее удобными для этой цели являются комплексы фиксированных действий (КФД) благодаря их четкой выраженности и устойчивости.
В. С. Дилгер (1962) наблюдал гнездостроительное поведение у разных видов попугаев-неразлучников. Особи одного вида отрывали кусочки материала для гнезда и переносили их в клюве, а особи другого не держали их в клюве, а засовывали под боковые перья. Гибриды, полученные от скрещивания этих видов, обнаружили смешанный тип поведения: птицы вначале засовывали материал для гнезда в перья, а потом вынимали его и брали в клюв, доставляя к гнезду. Такая смесь двух КФД была представлена у межвидовых гибридов. Следовательно, у гибридов в их генотипе представлены оба родительских поведенческих комплекса, однако извращена целесообразная последовательность их реализации.
Имеется много примеров, показывающих существование генов, действие которых подобно переключателю: они определяют, появится ли данный комплекс действия или нет. Вероятно, естественный отбор благоприятствует группировке тех генов, которые способствуют проявлению полезного поведенческого признака.
Эффективным путем для изучения эволюции поведения является вышеописанный прием экспериментального анализа наследования КФД. Метод получения межвидовых гибридов оказывается в этом плане весьма убедительным.
Нервно-психические заболевания генетической природы. К настоящему времени известно более 80 заболеваний, связанных с определенными генетическими дефектами. И хотя очевидно, что наследственная информация влияет на развитие психопатологических симптомов, конкретные механизмы этого влияния в большинстве случаев остаются неизвестными.
Определенные виды расстройств высшей нервной деятельности связывают с тремя типами генетических аномалий.
1. Аномалии, связанные с рецессивными генами, возникают в результате дефицита определенного фермента в печени. Типичным примером такого рода заболевания служит фенилкетонурия, которая в поведении проявляется в форме задержки умственного развития, а иногда сопровождается эпилептическими припадками и психозами. Данное заболевание обусловлено изменением генетического кода в одном участке хромосомы.
2. Аномалии, связанные с доминантными генами, в заболевании проявляются при особых условиях или стрессах. Ярким примером может служить хорея Геттингтона, которая начинается обычно в возрасте 30—40 лет (дегенерация коры и базальных ганглиев) и проявляется в непроизвольных движениях, неправильной подергивающейся походке, характерных гримасах, замедленной, невнятной речи, ухудшении памяти, повышенной раздражительности, депрессии, слабоумии.
Сходны с этой болезнью и такие наследственные заболевания нервной системы, как болезнь Альцгеймера и болезнь Паркинсона (дрожательный паралич). Допуска-
ют, что такую же природу имеют шизофрения (грубое расстройство интеллектуальной деятельности) и маниакально-депрессивные психозы. Высокая наследственная предрасположенность к шизофрении не вызывает сомнения. Риск заболевания шизофренией у детей, где один из родителей болен шизофренией, 10-15 и 35-40%, если больны оба родителя. Существуют данные о высокой наследственной предрасположенности к эпилепсии, основным симптомом которой являются судорожные припадки.
3. Аномалии, связанные с хромосомными аберрациями, заключающимися в добавлении или утрате целой хромосомы или ее части. Типичным примером такого заболевания служит болезнь Дауна, выражающаяся в задержке умственного и психического развития. Больной имеет 47 хромосом вместо 46, свойственных нормальному человеку. Наличие лишней хромосомы обусловливает синтез избыточного количества фермента, необходимого для построения белков головного мозга.
Группу заболеваний с ранним нарушением интеллекта объединяют под термином олигофрения. При этом хромосомные мутации могут выражаться в изменении числа хромосом, их структуры или краткого изменения гаплоидного набора. И всегда они сопровождаются тяжелыми формами умственной отсталости в сочетании с различными пороками развития.
Л. Г. Романова и Л. Г. Калмыкова (1981)выдвинули гипотезу о генетической природе психических болезней. Они полагают, что доминантные мутации должны приводить к поражению или дисфункции строго ограниченных структур мозга. Приуроченность данного психического дефекта к определенным жестким мозговым системам дает возможность понять не только мозговые механизмы психических расстройств, но и определенные аспекты нормального поведения.
Генетика свойств нервных процессов. Первым, кто обратил внимание на необходимость изучить генетическую природу свойств основных нервных процессов (их силы, уравновешенности и подвижности), был И. П. Павлов в процессе разработки им физиологической базы для определения типов нервной системы животных.
В дальнейшем отечественные ученые (Л. В. Кру-шинский, 1946; В. К. Красусский, 1963) доказали генетическую детерминированность силы возбудительного процесса, а В. К. Федоров (1971) показал зависимость от генотипа материнского организма такого свойства, как подвижность нервных процессов.
М. Е. Лобашев обратил внимание на наследование такого фундаментального свойства нервной системы, как возбудимость, которая определяет действие генов, детерминирующих отдельные поведенческие акты.
Ученики М. Е. Лобашева создали стройную систему взглядов на нейрохимическую и молекулярную детерминацию поведения (Н. Г. Лопатина, В. В. Пономарен-ко, 1987).
Исследования позволили вычленить эффект отдельного гена, вносящего основной вклад в формирование порога нервно-мышечной возбудимости, а также установить его идентичность с определенным локусом хромосомы. Более того, эти же гены участвуют в наследственной детерминации пороговых характеристик нервной системы в целом. У видов, пород и рас животных, имеющих высокую нервно-мышечную возбудимость, наблюдали и более высокую пищевую возбудимость и более высокие показатели силы возбуждения. Основой такой общности генетических механизмов могут быть функциональные или структурные особенности клеточных мембран и связанных с ними ферментов, определяющих характер протекания внутриклеточных процессов (В. В. Пономаренко, 1970).
Генетическая детерминация особенностей обучения. Выведены линии крыс, различающихся по обучаемости в сложном лабиринте: способные крысы ориентировались лучше по пространственным ориентирам, а неспособные — по зрительным. Большое значение при этом имеет характер мотивации: способные крысы сильнее мотивируются голодом, а неспособные — в защитно-оборонительных ситуациях.
Вклад каждой из скрещиваемых линий в свойства обучаемости конкретного поколения и особи неодинаков. М. П. Садовникова-Кольцова (1928) предположила наличие у крыс трех групп генов, детерминирующих способ-
ность к обучению: гены двигательной активности, гены эмоции страха и гены ориентировочно исследовательского инстинкта. Однако корреляция этих свойств со способностью к обучению не всегда проявляется, и в настоящее время рассматривают эти комбинации признаков как носящие фенотипический характер.
Наследственную основу различий в обучении М. Е. Лобашев предполагал искать в генетически детерминированных особенностях безусловных рефлексов, и прежде всего в уровне пищевого рефлекса как основного модулятора функциональной активности нервной системы. Показано наличие генетических корреляций между уровнем функциональной активности нервной системы и способностью к выработке условных рефлексов (В. В. Пономаренко).
Ю. С. Дмитриевым (1981) было установлено, что порог возбудимости и скорость образования оборонительных условных рефлексов проявляют сходный характер наследования (рис. 25). Вероятно, в основе лежит мно жественный эффект какой-то общей группы генов.
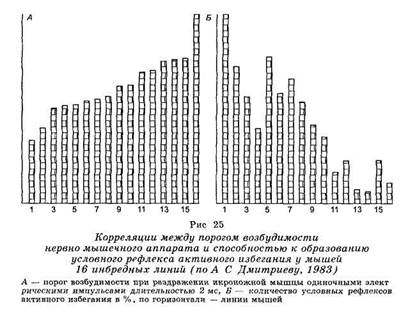
В число контролируемых им признаков поведения следует отнести ориентировочно-исследовательскую активность, общую двигательную активность и уровень эмоционального статуса. Последние могут участвовать в наследственной обусловленности условнорефлекторных форм поведения. Ген, контролирующий вышеназванные признаки, идентичен гену, определяющему содержание биологически активных соединений в плазме крови, серотонина и норадреналина в тканях гипоталамуса (А. Оливерио и др., 1973; Ю. С. Дмитриев, 1981).
Таким образом, множественное влияние одного и того же гена может заключаться в контроле порога возбудимости нервной системы, содержания нейроактивных соединений и способности к обучению (образованию оборонительных условных рефлексов).
Генотип и анатомические особенности мозга. Благодаря трансплантации появилась возможность экспериментального изменения массы мозга. Пересадка африканской рыбке-теляпии среднего мозга от донора увеличила общую массу мозга реципиента и одновременно улучшила ее интеллектуальные способности: рыбка успешно выполняла задачи по различению и переделке сигнального значения раздражителей, что до операции делала с трудом.
К настоящему времени получено значительное число убедительных аргументов, которые позволили приписывать особую роль структур

