 2013-12-31
2013-12-31 1256
1256Чтобы понять сущность процессов обмена энергии, необходимо познакомиться с рядом общих положений термодинамики. Первый закон термодинамики – это закон о сохранении энергии. Этот закон утверждает, что энергию нельзя ни создать, ни уничтожить. Математически первый закон термодинамики можно выразить формулой:
Q = ∆H + W',
где Q – энергия в виде теплоты; ∆H – изменение энтальпии (или теплосодержания системы) при постоянном давлении; W' – полезная работа, выполненная благодаря введению определенного количества теплоты Q.
Данное уравнение справедливо для идеальной обратимой системы, т.е. такой системы, которая, будучи приведенной, в новое состояние путем поглощения энергии, может вновь возвратиться в исходное состояние и освободить то же самое количество ранее поглощенной энергии. Последняя может быть использована для совершения новой работы. Однако в природе полностью обратимых систем не существует. Самопроизвольно химические процессы могут протекать лишь в одном направлении – в направлении состояния равновесия. Предсказать направление биохимических реакций позволяет второй закон термодинамики, согласно которому все самопроизвольные процессы протекают в направлении, соответствующем максимальной при данных условиях энтропии (S), до достижения состояния равновесия системы. Энтропия (S) – мера разупорядоченности системы. Увеличение энтропии системы препятствует возврату к исходному состоянию. Поэтому все реакции, которые сопровождаются возрастанием энтропии, необратимы. Для обращения их затрачивают дополнительную энергию.
Для осуществления катаболизма и анаболизма клеткам необходима свободная энергия, поскольку только свободная энергия способна производить работу при постоянной температуре и давлении. Ее изменение обозначают символом ∆G. Ту же часть энергии химического процесса, которая не может быть превращена в работу, называют связанной энергией и выражают произведением T∆S (где T – абсолютная температура, а ∆S – изменение энтропии системы при данном химическом превращении). Сумма изменений свободной и связанной энергий называется энтальпией (или теплосодержанием системы). И, как уже отмечалось, обозначается символом ∆H. Второй закон термодинамики можно выразить формулой:
∆H = ∆G + T∆S или ∆G = ∆H – T∆S
Если (например, в ходе реакции А → В) величина ∆G имеет отрицательное значение, то это означает, что свободная энергия вещества В меньше, чем свободная энергия вещества А. в этом случае реакция может происходить самопроизвольно с выделением свободной энергии (экзотермический процесс). Наоборот, если ∆G – положительная величина, то свободная энергия у вещества В выше, чем у соединения А, и самопроизвольно реакция протекать не может. Она протекает с потреблением энергии извне (эндотермический процесс). Если система находится в состоянии равновесия, то количество свободной энергии (∆G) равно нулю.
Величины ∆G могут выражаться в джоулях на моль или калориях на моль.
Изменение стандартной свободной энергии химической реакции можно вычислить:
∆Go = – RT lnК,
где К – константа равновесия, R – универсальная газовая постоянная (1,987 кал/моль*К).
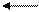
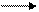 А + В С + Д,
А + В С + Д,
 К = [C] [Д]
К = [C] [Д]
[A] [B],
Стандартные условия: t = 25 oC, давление 1 атм, [A], [B], [C], [Д] – молярная концентрация реагирующих веществ в состоянии равновесия при стандартных условиях. Если К > 1, то ∆Go > 0; при К < 1, ∆Go < 0.
∆Go – это разность между свободной энергией исходных веществ и свободной энергией продуктов реакции при стандартных условиях и исходной концентрации веществ 1 М.
Отрицательное значение ∆Go означает, что в продуктах реакции содержится меньше свободной энергии, чем в исходных веществах. Поскольку реакции идут в направлении, соответствующем уменьшению свободной энергии, то в этом случае равновесие будет смещено вправо.
Положительное значение ∆Go означает, что продукты реакции содержат больше свободной энергии, чем исходные вещества.
Биохимические реакции происходят обычно при рН = 7,0, потому в биоэнергетике в качестве стандартного принято состояние при рН = 7,0. Изменение свободной стандартной энергии обозначается при этом ∆Go '.
Пример. Вычислим ∆G'стандартной свободной энергии для реакции, катализируемой ферментом фосфоглюкомутазой.
ФГМ
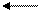
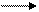 Гл-1-Р Гл-6-Р
Гл-1-Р Гл-6-Р
Фермент не смещает равновесие реакций, а только ускоряет достижение равновесия. Измеренные равновесные концентрации
Гл-1-Р – 0,001 М
Гл-6-Р – 0,019 М
К' = 0,019 / 0,001 = 19
∆Go ' = – 1,7 ккал / моль,
Таким образом, превращение Гл-1-Р в Гл-6-Р протекает с потерей свободной энергии.
Величина ∆Go ' – строго определенная константа, характерная для каждой реакции, причем, в стандартных условиях 1 М; рН = 7,0; t = 25 оС.
Истинное же изменение свободной энергии ∆G для данной химической реакции зависит от условий, при которых эта реакция фактически протекает (условия – отличные от стандартных). ∆G связана с величиной ∆Go ' выражением
∆G = ∆Go ' + RT ln К
Пример. В эритроцитах человека установлены равновесные концентрации АТФ – 2,25 мМ; АДФ – 0,25 мМ; Рн – 1,65 мМ. Примем для простоты что t = 25 оС, рН = 7,0.
Истинное изменение свободной энергии гидролиза:
∆G = ∆Go ' + RT ln К =
= – 7300 + 1360 lg [ 2,5· 10-4 · 1,65·10-3]: 2,25·10-3 =
= – 7300 + 1300 lg 1,83 ·10-4 = –12,4 ккал / моль.
При синтезе АТФ из АДФ и Рн – затрачивается 12,4 ккал / моль.
Изменения стандартной свободной энергии последовательно протекающих химических реакций аддитивны, т.е.
∆Go ' 3= ∆Go ' 1+ ∆Go '2
АТФ – главный химический посредник клетки, связывающий между собой процессы, идущие с выделением или потреблением энергии.
Характерное значение стандартной свободной энергии гидролиза АТФ можно вычислить. В стандартных условиях для реакции

 АТФ + Н2О АДФ + Рн
АТФ + Н2О АДФ + Рн
∆Go ' составляет – 7,3 ккал / моль, ∆Go ' для реакции

 АДФ + Н2О АМФ + Рн
АДФ + Н2О АМФ + Рн
также составляет – 7,3 ккал / моль.
Величина ∆Go ' гидролиза АМФ уже значительно ниже – 3,4 ккал / моль.
Хотя при стандартных условиях величина ∆Go ' гидролиза АТФ – 7,3 ккал / моль, истинное значение в интактных клетках может отличаться от этой величины. Объясняется это тем, что концентрации АТФ, АДФ и Рн в живых клетках неодинаковы, намного ниже стандартной концентрации 1М.
Высокоэнергетические фосфаты. Освобождающаяся в клетках в результате катаболических процессов распада питательных веществ свободная энергия в дальнейшем может быть использована для осуществления многих химических реакций, протекающих с затратой энергии.
Запасание энергии происходит в виде богатых энергией химических связей особого класса соединений, большинство из которых является ангидридами фосфорной кислоты (нуклеозидтрифосфаты).
Существуют высокоэнергетические и низкоэнергетические фосфаты. Условной границей для этих двух групп соединений является величина свободной энергии гидролиза фосфатной связи. Следовательно, высокоэнергетические фосфаты имеют богатую энергией высокоэргическую (макроэргическую) фосфатную связь. Когда говорят о богатых энергиях связях, то в этом случае энергию связи определяют как разницу свободных энергий соединений, содержащего эту связь, и соединений, получающихся после ее разрыва. Макроэргическими (богатыми энергией) принято считать те связи, при гидролизе которых изменения свободной энергии системы (–∆G) составляют более 21 кДж/моль (или более 5 ккал/моль).
Центральную роль в энергообмене клеток всех типов осуществляет система адениновых нуклеотидов, которая включает в себя АТФ, АДФ и АМФ, а также неорганический фосфат (Фн) и ионы магния. АТФ является термодинамически неустойчивой молекулой и гидролизуется с образованием АДФ или АМФ. Именно эта неустойчивость позволяет АТФ выполнять функцию переносчика химической энергии, необходимой для удовлетворения большей части энергетических потребностей клеток. К соединениям, обладающим богатой энергией связью, помимо АТФ, относятся также УТФ, ЦТФ, ГТФ, ТТФ, креатин фосфат, пирофосфат, некоторые тиоэфиры (например, ацетил-КоА), фосфоенолпируват, 1,3-бисфосфоглицерат и ряд других соединений. Однако образование этих соединений в большинстве случаев зависит от энергии, поставляемой АТФ.
Стандартные условия: 1,0 М концентрации исходных и конечных продуктов, рН 7,0, температура 37оС и избыток ионов магния. В этих условиях при гидролизе АТФ (АТФ + Н2О → АДФ + Фн) изменение свободной энергии (–∆G) составляет –30,4 кДж/моль. При физиологических условиях в клетке не имеется таких высоких концентраций веществ и их продуктов, а также ионов магния. Кроме того, возможны отклонения и в значениях рН. Поэтому в физиологических условиях реальная свободная энергия (–∆Gф) гидролиза концевой фосфатной связи АТФ будет иная и приближается к –50,0 кДж/моль.
Величина –∆G для других соединений отличается от стандартной (–∆Gо), но не обязательно в сторону больших значений.
Заметим, что АТФ, хотя и служит ключевым энергетическим посредником в обмене веществ, не является веществом наиболее «богатым» энергией. АТФ находится в середине энергетической шкалы.
Возможно несколько вариантов освобождения энергии фосфатных связей АТФ. Основной вариант – это отщепление концевого фосфата АТФ (АТФ + Н2О → АДФ + Н3РО4). Этот тип реакций значительно реже используется в биохимических процессах. Образующийся пирофосфат относится к «богатым энергией» веществам. Однако богатые энергией связи пирофосфата редко используются для синтеза других веществ, т.к. при его гидролизе энергия освобождается в виде теплоты.
При гидролизе концевой фосфатной связи АДФ высвобождается такое же количество энергии, как и при отщеплении концевой фосфатной связи АТФ. Казалось бы, что АДФ может во многих реакциях заменить АТФ, в частности в реакциях фосфорилирования. Однако эта возможность не реализуется в биохимических процессах. До сих пор реакции фосфорилирования при участии АДФ неизвестны. В действительности происходит гидролиз АДФ (АДФ + Н2О → АМФ + Н3РО4) до низкоэнергетических АМФ и фосфата с выделением теплоты.
Заключение. Метаболизм – это закономерный порядок превращения веществ и энергии в живых системах, направленный на их сохранение и самовоспроизведение. Обмен веществ и энергии неразрывно связаны. Первичным источником энергии для всей живой природы является солнечное излучение. Автотрофные и гетеротрофные организмы. Три главные стадии в аэробном катаболизме. Биоэнергетика клетки. Свободная энергия. Накопление энергии в специфических фосфатных связях АТФ лежит в основе механизма переноса энергии в живой клетке. Три основных типа перехода энергии АТФ в клетке: в энергию химических связей, в тепловую энергию и энергию, затрачиваемую на совершение работы (осмотической, электрической, механической и др.).








