 2014-02-09
2014-02-09 9395
9395Мейоз
Мейоз — это особый способ деления эукариотических клеток, в результате которого происходит переход клеток из диплоидного состояния в гаплоидное. Мейоз состоит из двух последовательных делений, которым предшествует однократная репликация ДНК.
Первое мейотическое деление (мейоз 1) называется редукционным, поскольку именно во время этого деления происходит уменьшение числа хромосом вдвое: из одной диплоидной клетки (2n 4c) образуются две гаплоидные (1n 2c).
Интерфаза 1 (в начале — 2n 2c, в конце — 2n 4c) — синтез и накопление веществ и энергии, необходимых для осуществления обоих делений, увеличение размеров клетки и числа органоидов, удвоение центриолей, репликация ДНК, которая завершается в профазе 1.
Профаза 1 (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом, конъюгация гомологичных хромосом и кроссинговер. Конъюгация — процесс сближения и переплетения гомологичных хромосом. Пару конъюгирующих гомологичных хромосом называют бивалентом. Кроссинговер — процесс обмена гомологичными участками между гомологичными хромосомами.
Профаза 1 подразделяется на стадии: лептотена (завершение репликации ДНК), зиготена (конъюгация гомологичных хромосом, образование бивалентов), пахитена (кроссинговер, перекомбинация генов), диплотена (выявление хиазм, 1 блок овогенеза у человека), диакинез (терминализация хиазм).
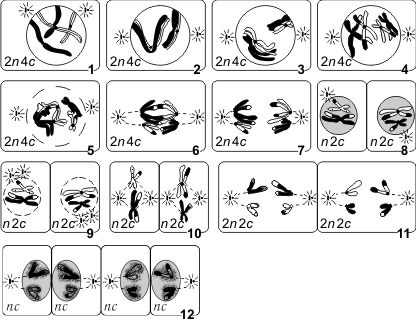
Мейоз: 1 — лептотена; 2 — зиготена; 3 — пахитена; 4 — диплотена; 5 — диакинез; 6 — метафаза 1; 7 — анафаза 1; 8 — телофаза 1; 9 — профаза 2; 10 — метафаза 2; 11 — анафаза 2; 12 — телофаза 2.
Метафаза 1 (2n 4c) — выстраивание бивалентов в экваториальной плоскости клетки, прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза 1 (2n 4c) — случайное независимое расхождение двухроматидных хромосом к противоположным полюсам клетки (из каждой пары гомологичных хромосом одна хромосома отходит к одному полюсу, другая — к другому), перекомбинация хромосом.
Телофаза 1 (1n 2c в каждой клетке) — образование ядерных мембран вокруг групп двухроматидных хромосом, деление цитоплазмы. У многих растений клетка из анафазы 1 сразу же переходит в профазу 2.
Второе мейотическое деление (мейоз 2) называется эквационным.
Интерфаза 2, или интеркинез (1n 2c), представляет собой короткий перерыв между первым и вторым мейотическими делениями, во время которого не происходит репликация ДНК. Характерна для животных клеток.
Профаза 2 (1n 2c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления.
Метафаза 2 (1n 2c) — выстраивание двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом; 2 блок овогенеза у человека.
Анафаза 2 (2n 2с) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами), перекомбинация хромосом.
Телофаза 2 (1n 1c в каждой клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия) с образованием в итоге четырех гаплоидных клеток.
Биологическое значение мейоза. Мейоз является центральным событием гаметогенеза у животных и спорогенеза у растений. Являясь основой комбинативной изменчивости, мейоз обеспечивает генетическое разнообразие гамет.
Амитоз
Амитоз — прямое деление интерфазного ядра путем перетяжки без образования хромосом, вне митотического цикла. Описан для стареющих, патологически измененных и обреченных на гибель клеток. После амитоза клетка не способна вернуться в нормальный митотический цикл.
Клеточный цикл
Клеточный цикл — жизнь клетки от момента ее появления до деления или смерти. Обязательным компонентом клеточного цикла является митотический цикл, который включает в себя период подготовки к делению и собственно митоз. Кроме этого, в жизненном цикле имеются периоды покоя, во время которых клетка выполняет свойственные ей функции и избирает дальнейшую судьбу: гибель или возврат в митотический цикл.
Размножение организмов
Размножение — свойство живых организмов воспроизводить себе подобных. Существуют два основных способа размножения — бесполое и половое.
Бесполое размножение
Бесполое размножение осуществляется при участии лишь одной родительской особи и происходит без образования гамет. Дочернее поколение у одних видов возникает из одной или группы клеток материнского организма, у других видов — в специализированных органах. Различают следующие способы бесполого размножения: деление, почкование, фрагментация, полиэмбриония, спорообразование, вегетативное размножение.
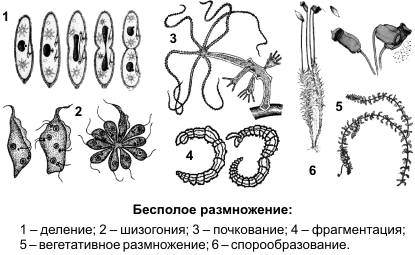
Деление — способ бесполого размножения, характерный для одноклеточных организмов, при котором материнская особь делится на две или большее количество дочерних клеток.
Можно выделить:
а) простое бинарное деление (прокариоты),
б) митотическое бинарное деление (простейшие, одноклеточные водоросли),
в) множественное деление, или шизогонию (малярийный плазмодий, трипаносомы).
Во время деления парамеции (1) микронуклеус делится митозом, макронуклеус — амитозом. Во время шизогонии (2) сперва многократно митозом делится ядро, затем каждое из дочерних ядер окружается цитоплазмой, и формируются несколько самостоятельных организмов.
Почкование — способ бесполого размножения, при котором новые особи образуются в виде выростов на теле родительской особи (3). Дочерние особи могут отделяться от материнской и переходить к самостоятельному образу жизни (гидра, дрожжи), могут остаться прикрепленными к ней, образуя в этом случае колонии (коралловые полипы).
Фрагментация (4) — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается материнская особь (кольчатые черви, морские звезды, спирогира, элодея). В основе фрагментации лежит способность организмов к регенерации.
Полиэмбриония — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается эмбрион (монозиготные близнецы).
Вегетативное размножение — способ бесполого размножения, при котором новые особи образуются или из частей вегетативного тела материнской особи, или из особых структур (корневище, клубень и др.), специально предназначенных для этой формы размножения. Вегетативное размножение характерно для многих групп растений, используется в садоводстве, огородничестве, селекции растений (искусственное вегетативное размножение).
| Вегетативный орган | Способ вегетативного размножения | Примеры |
| Корень | Корневые черенки | Шиповник, малина, осина, ива, одуванчик |
| Корневые отпрыски | Вишня, слива, осот, бодяк, сирень | |
| Надземные части побегов | Деление кустов | Флокс, маргаритка, примула, ревень |
| Стеблевые черенки | Виноград, смородина, крыжовник | |
| Отводки | Крыжовник, виноград, черемуха | |
| Подземные части побегов | Корневище | Спаржа, бамбук, ирис, ландыш |
| Клубень | Картофель, седмичник, топинамбур | |
| Луковица | Лук, чеснок, тюльпан, гиацинт | |
| Клубнелуковица | Гладиолус, крокус | |
| Лист | Листовые черенки | Бегония, глоксиния, колеус |
Спорообразование (6) — размножение посредством спор. Споры — специализированные клетки, у большинства видов образуются в особых органах — спорангиях. У высших растений образованию спор предшествует мейоз.
Клонирование — комплекс методов, используемых человеком для получения генетически идентичных копий клеток или особей. Клон — совокупность клеток или особей, произошедших от общего предка путем бесполого размножения. В основе получения клона лежит митоз (у бактерий — простое деление).
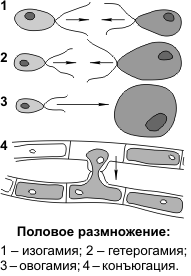
Половое размножение осуществляется при участии двух родительских особей (мужской и женской), у которых в особых органах образуются специализированные клетки — гаметы. Процесс формирования гамет называется гаметогенезом, основным этапом гаметогенеза является мейоз. Дочернее поколение развивается из зиготы — клетки, образовавшейся в результате слияния мужской и женской гамет. Процесс слияния мужской и женской гамет называется оплодотворением. Обязательным следствием полового размножения является перекомбинация генетического материала у дочернего поколения.
В зависимости от особенностей строения гамет, можно выделить следующие формы полового размножения: изогамию, гетерогамию и овогамию.
Изогамия (1) — форма полового размножения, при которой гаметы (условно женские и условно мужские) являются подвижными и имеют одинаковые морфологию и размеры.
Гетерогамия (2) — форма полового размножения, при которой женские и мужские гаметы являются подвижными, но женские — крупнее мужских и менее подвижны.
Овогамия (3) — форма полового размножения, при которой женские гаметы неподвижные и более крупные, чем мужские гаметы. В этом случае женские гаметы называются яйцеклетками, мужские гаметы, если имеют жгутики, — сперматозоидами, если не имеют, — спермиями.
Овогамия характерна для большинства видов животных и растений. Изогамия и гетерогамия встречаются у некоторых примитивных организмов (водоросли). Кроме вышеперечисленных, у некоторых водорослей и грибов имеются формы размножения, при которых половые клетки не образуются: хологамия и конъюгация. При хологамии происходит слияние друг с другом одноклеточных гаплоидных организмов, которые в данном случае выступают в роли гамет. Образовавшаяся диплоидная зигота затем делится мейозом с образованием четырех гаплоидных организмов. При конъюгации (4) происходит слияние содержимого отдельных гаплоидных клеток нитевидных талломов. По специально образующимся каналам содержимое одной клетки перетекает в другую, образуется диплоидная зигота, которая обычно после периода покоя также делится мейозом.
Основные понятия генетики. Законы Менделя
Генетика — наука о закономерностях наследственности и изменчивости. Датой «рождения» генетики можно считать 1900 год, когда Г. Де Фриз в Голландии, К. Корренс в Германии и Э. Чермак в Австрии независимо друг от друга «переоткрыли» законы наследования признаков, установленные Г. Менделем еще в 1865 году.
Наследственность — свойство организмов передавать свои признаки от одного поколения к другому.
Изменчивость — свойство организмов приобретать новые по сравнению с родителями признаки. В широком смысле под изменчивостью понимают различия между особями одного вида.
Признак — любая особенность строения, любое свойство организма. Развитие признака зависит как от присутствия других генов, так и от условий среды, формирование признаков происходит в ходе индивидуального развития особей. Поэтому каждая отдельно взятая особь обладает набором признаков, характерных только для нее.
Фенотип — совокупность всех внешних и внутренних признаков организма.
Ген — функционально неделимая единица генетического материала, участок молекулы ДНК, кодирующий первичную структуру полипептида, молекулы транспортной или рибосомной РНК. В широком смысле ген — участок ДНК, определяющий возможность развития отдельного элементарного признака.
Генотип — совокупность генов организма.
Локус — местоположение гена в хромосоме.
Аллельные гены — гены, расположенные в идентичных локусах гомологичных хромосом.
Гомозигота — организм, имеющий аллельные гены одной молекулярной формы.
Гетерозигота — организм, имеющий аллельные гены разной молекулярной формы; в этом случае один из генов является доминантным, другой — рецессивным.
Рецессивный ген — аллель, определяющий развитие признака только в гомозиготном состоянии; такой признак будет называться рецессивным.
Доминантный ген — аллель, определяющий развитие признака не только в гомозиготном, но и в гетерозиготном состоянии; такой признак будет называться доминантным.
Методы генетики
Основным является гибридологический метод — система скрещиваний, позволяющая проследить закономерности наследования признаков в ряду поколений. Впервые разработан и использован Г. Менделем.
Отличительные особенности метода:
1) целенаправленный подбор родителей, различающихся по одной, двум, трем и т. д. парам контрастных (альтернативных) стабильных признаков;
2) строгий количественный учет наследования признаков у гибридов;
3) индивидуальная оценка потомства от каждого родителя в ряду поколений.
Скрещивание, при котором анализируется наследование одной пары альтернативных признаков, называется моногибридным, двух пар — дигибридным, нескольких пар — полигибридным. Под альтернативными признаками понимаются различные значения какого-либо признака, например, признак — цвет горошин, альтернативные признаки — желтый цвет, зеленый цвет горошин.
Кроме гибридологического метода, в генетике используют: генеалогический — составление и анализ родословных; цитогенетический — изучение хромосом; близнецовый — изучение близнецов; популяционно-статистический метод — изучение генетической структуры популяций.
Генетическая символика
Предложена Г. Менделем, используется для записи результатов скрещиваний: Р — родители; F — потомство, число внизу или сразу после буквы указывает на порядковый номер поколения (F1 — гибриды первого поколения — прямые потомки родителей, F2 — гибриды второго поколения — возникают в результате скрещивания между собой гибридов F1); × — значок скрещивания; G — мужская особь; E — женская особь; A — доминантный ген, а — рецессивный ген; АА — гомозигота по доминанте, аа — гомозигота по рецессиву, Аа — гетерозигота.
Закон единообразия гибридов первого поколения, или первый закон Менделя
Успеху работы Менделя способствовал удачный выбор объекта для проведения скрещиваний — различные сорта гороха.
Особенности гороха:
1) относительно просто выращивается и имеет короткий период развития;
2) имеет многочисленное потомство;
3) имеет большое количество хорошо заметных альтернативных признаков (окраска венчика — белая или красная; окраска семядолей — зеленая или желтая; форма семени — морщинистая или гладкая; окраска боба — желтая или зеленая; форма боба — округлая или с перетяжками; расположение цветков или плодов — по всей длине стебля или у его верхушки; высота стебля — длинный или короткий);
4) является самоопылителем, в результате чего имеет большое количество чистых линий, устойчиво сохраняющих свои признаки из поколения в поколение.
Опыты по скрещиванию разных сортов гороха Мендель проводил в течение восьми лет, начиная с 1854 года. 8 февраля 1865 года Г. Мендель выступил на заседании Брюннского общества естествоиспытателей с докладом «Опыты над растительными гибридами», где были обобщены результаты его работы.
Опыты Менделя были тщательно продуманы. Если его предшественники пытались изучить закономерности наследования сразу многих признаков, то Мендель свои исследования начал с изучения наследования всего лишь одной пары альтернативных признаков.
Мендель взял сорта гороха с желтыми и зелеными семенами и произвел их искусственное перекрестное опыление: у одного сорта удалил тычинки и опылил их пыльцой другого сорта. Гибриды первого поколения имели желтые семена. Аналогичная картина наблюдалась и при скрещиваниях, в которых изучалось наследование других признаков: при скрещивании растений, имеющих гладкую и морщинистую формы семян, все семена полученных гибридов были гладкими, от скрещивания красноцветковых растений с белоцветковыми все полученные — красноцветковые. Мендель пришел к выводу, что у гибридов первого поколения из каждой пары альтернативных признаков проявляется только один, а второй как бы исчезает. Проявляющийся у гибридов первого поколения признак Мендель назвал доминантным, а подавляемый — рецессивным.
При моногибридном скрещивании гомозиготных особей, имеющих разные значения альтернативных признаков, гибриды являются единообразными по генотипу и фенотипу.
Генетическая схема закона единообразия Менделя
(А — желтый цвет горошин, а — зеленый цвет горошин)
| Р | ?AA желтые | × | ?аа зеленые |
| Типы гамет |  А А | а | |
| F1 | Aа желтые 100% |
Закон расщепления, или второй закон Менделя
Г. Мендель дал возможность самоопылиться гибридам первого поколения. У полученных таким образом гибридов второго поколения проявился не только доминантный, но и рецессивный признак. Результаты опытов приведены в таблице.
| Признаки | Доминантные | Рецессивные | Всего | ||
| Число | % | Число | % | ||
| Форма семян | 74,74 | 25,26 | |||
| Окраска семядолей | 75,06 | 24,94 | |||
| Окраска семенной кожуры | 75,90 | 24,10 | |||
| Форма боба | 74,68 | 25,32 | |||
| Окраска боба | 73,79 | 26,21 | |||
| Расположение цветков | 75,87 | 24,13 | |||
| Высота стебля | 73,96 | 26,04 | |||
| Всего: | 74,90 | 25,10 |
Анализ данных таблицы позволил сделать следующие выводы:
- единообразия гибридов во втором поколении не наблюдается: часть гибридов несет один (доминантный), часть — другой (рецессивный) признак из альтернативной пары;
- количество гибридов, несущих доминантный признак, приблизительно в три раза больше, чем гибридов, несущих рецессивный признак;
- рецессивный признак у гибридов первого поколения не исчезает, а лишь подавляется и проявляется во втором гибридном поколении.
Явление, при котором часть гибридов второго поколения несет доминантный признак, а часть — рецессивный, называют расщеплением. Причем, наблюдающееся у гибридов расщепление не случайное, а подчиняется определенным количественным закономерностям. На основе этого Мендель сделал еще один вывод: при скрещивании гибридов первого поколения в потомстве происходит расщепление признаков в определенном числовом соотношении.
При моногибридном скрещивании гетерозиготных особей у гибридов имеет место расщепление по фенотипу в отношении 3:1, по генотипу 1:2:1.
Генетическая схема закона расщепления Менделя
(А — желтый цвет горошин, а — зеленый цвет горошин):
| P | ?Aa желтые | × | ?Aa желтые | ||
| Типы гамет | A | a | A | a | |
| F2 | AA желтые | Aa желтые 75% | Aa желтые | aa зеленые 25% |
С 1854 года в течение восьми лет Мендель проводил опыты по скрещиванию растений гороха. Им было выявлено, что в результате скрещивания различных сортов гороха друг с другом гибриды первого поколения обладают одинаковым фенотипом, а у гибридов второго поколения имеет место расщепление признаков в определенных соотношениях. Для объяснения этого явления Мендель сделал ряд предположений, которые получили название «гипотезы чистоты гамет», или «закона чистоты гамет».
Мендель предположил, что:
· за формирование признаков отвечают какие-то дискретные наследственные факторы;
· организмы содержат два фактора, определяющих развитие признака;
· при образовании гамет в каждую из них попадает только один из пары факторов;
· при слиянии мужской и женской гамет эти наследственные факторы не смешиваются (остаются чистыми).
В 1909 году В. Иогансен назовет эти наследственные факторы генами, а в 1912 году Т. Морган покажет, что они находятся в хромосомах.
Для доказательства своих предположений Г. Мендель использовал скрещивание, которое сейчас называют анализирующим (анализирующее скрещивание — скрещивание организма, имеющего неизвестный генотип, с организмом, гомозиготным по рецессиву). Наверное, Мендель рассуждал следующим образом: «Если мои предположения верны, то в результате скрещивания F1 с сортом, обладающим рецессивным признаком (зелеными горошинами), среди гибридов будут половина горошин зеленого цвета и половина горошин — желтого». Как видно из приведенной ниже генетической схемы, он действительно получил расщепление 1:1 и убедился в правильности своих предположений и выводов, но современниками он понят не был. Его доклад «Опыты над растительными гибридами», сделанный на заседании Брюннского общества естествоиспытателей, был встречен полным молчанием.
| Р | ?Аа желтые | × | ?aа зеленые |
| Типы гамет | A a | a | |
| F | Аа желтые 50% | аa зеленые 50% |
Цитологические основы первого и второго законов Менделя
Во времена Менделя строение и развитие половых клеток не было изучено, поэтому его гипотеза чистоты гамет является примером гениального предвидения, которое позже нашло научное подтверждение.
Явления доминирования и расщепления признаков, наблюдавшиеся Менделем, в настоящее время объясняются парностью хромосом, расхождением хромосом во время мейоза и объединением их во время оплодотворения. Обозначим ген, определяющий желтую окраску, буквой А, а зеленую — а. Поскольку Мендель работал с чистыми линиями, оба скрещиваемых организма — гомозиготны, то есть несут два одинаковых аллеля гена окраски семян (соответственно, АА и аа). Во время мейоза число хромосом уменьшается в два раза, и в каждую гамету попадает только одна хромосома из пары. Так как гомологичные хромосомы несут одинаковые аллели, все гаметы одного организмы будут содержать хромосому с геном А, а другого — с геном а.
При оплодотворении мужская и женская гаметы сливаются, и их хромосомы объединяются в одной зиготе. Получившийся от скрещивания гибрид становится гетерозиготным, так как его клетки будут иметь генотип Аа; один вариант генотипа даст один вариант фенотипа — желтый цвет горошин.
У гибридного организма, имеющего генотип Аа во время мейоза, хромосомы расходятся в разные клетки и образуется два типа гамет — половина гамет будет нести ген А, другая половина — ген а. Оплодотворение — процесс случайный и равновероятный, то есть любой сперматозоид может оплодотворить любую яйцеклетку. Поскольку образовалось два типа сперматозоидов и два типа яйцеклеток, возможно возникновение четырех вариантов зигот. Половина из них — гетерозиготы (несут гены А и а), 1/4 — гомозиготы по доминантному признаку (несут два гена А) и 1/4 — гомозиготы по рецессивному признаку (несут два гена а). Гомозиготы по доминанте и гетерозиготы дадут горошины желтого цвета (3/4), гомозиготы по рецессиву — зеленого (1/4).
Закон независимого комбинирования (наследования) признаков, или третий закон Менделя
Организмы отличаются друг от друга по многим признакам. Поэтому, установив закономерности наследования одной пары признаков, Г. Мендель перешел к изучению наследования двух (и более) пар альтернативных признаков. Для дигибридного скрещивания Мендель брал гомозиготные растения гороха, отличающиеся по окраске семян (желтые и зеленые) и форме семян (гладкие и морщинистые). Желтая окраска (А) и гладкая форма (В) семян — доминантные признаки, зеленая окраска (а) и морщинистая форма (b) — рецессивные признаки.
Скрещивая растение с желтыми и гладкими семенами с растением с зелеными и морщинистыми семенами, Мендель получил единообразное гибридное поколение F1 с желтыми и гладкими семенами. От самоопыления 15-ти гибридов первого поколения было получено 556 семян, из них 315 желтых гладких, 101 желтое морщинистое, 108 зеленых гладких и 32 зеленых морщинистых (расщепление 9:3:3:1).
Анализируя полученное потомство, Мендель обратил внимание на то, что:
1) наряду с сочетаниями признаков исходных сортов (желтые гладкие и зеленые морщинистые семена), при дигибридном скрещивании появляются и новые сочетания признаков (желтые морщинистые и зеленые гладкие семена);
2) расщепление по каждому отдельно взятому признаку соответствует расщеплению при моногибридном скрещивании. Из 556 семян 423 были гладкими и 133 морщинистыми (соотношение 3:1), 416 семян имели желтую окраску, а 140 — зеленую (соотношение 3:1).
Мендель пришел к выводу, что расщепление по одной паре признаков не связано с расщеплением по другой паре. Для семян гибридов характерны не только сочетания признаков родительских растений (желтые гладкие семена и зеленые морщинистые семена), но и возникновение новых комбинаций признаков (желтые морщинистые семена и зеленые гладкие семена).
При дигибридном скрещивании дигетерозигот у гибридов имеет место расщепление по фенотипу в отношении 9:3:3:1, по генотипу в отношении 4:2:2:2:2:1:1:1:1, признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях.
| Р | ?АABB желтые, гладкие | × | ?aаbb зеленые, морщинистые |
| Типы гамет |  AB AB | ab | |
| F1 | AaBb желтые, гладкие, 100% | ||
| P | ?АaBb желтые, гладкие | × | ?AаBb желтые, гладкие |
| Типы гамет | AB Ab aB ab | AB Ab aB ab |
Генетическая схема закона независимого комбинирования признаков:
| Гаметы: | ? | AB | Ab | aB | ab |
| ? | |||||
| AB | AABB желтые гладкие | AABb желтые гладкие | AaBB желтые гладкие | AaBb желтые гладкие | |
| Ab | AABb желтые гладкие | AАbb желтые морщинистые | AaBb желтые гладкие | Aabb желтые морщинистые | |
| aB | AaBB желтые гладкие | AaBb желтые гладкие | aaBB зеленые гладкие | aaBb зеленые гладкие | |
| ab | AaBb желтые гладкие | Aabb желтые морщинистые | aaBb зеленые гладкие | aabb зеленые морщинистые |
Анализ результатов скрещивания по фенотипу: желтые, гладкие — 9/16, желтые, морщинистые — 3/16, зеленые, гладкие — 3/16, зеленые, морщинистые — 1/16. Расщепление по фенотипу 9:3:3:1.
Анализ результатов скрещивания по генотипу: AaBb — 4/16, AABb — 2/16, AaBB — 2/16, Aabb — 2/16, aaBb — 2/16, ААBB — 1/16, Aabb — 1/16, aaBB — 1/16, aabb — 1/16. Расщепление по генотипу 4:2:2:2:2:1:1:1:1.
Если при моногибридном скрещивании родительские организмы отличаются по одной паре признаков (желтые и зеленые семена) и дают во втором поколении два фенотипа (21) в соотношении (3 + 1)1, то при дигибридном они отличаются по двум парам признаков и дают во втором поколении четыре фенотипа (22) в соотношении (3 + 1)2. Легко посчитать, сколько фенотипов и в каком соотношении будет образовываться во втором поколении при тригибридном скрещивании: восемь фенотипов (23) в соотношении (3 + 1)3.
Если расщепление по генотипу в F2 при моногибридном поколении было 1:2:1, то есть было три разных генотипа (31), то при дигибридном образуется 9 разных генотипов — 32, при тригибридном скрещивании образуется 33 — 27 разных генотипов.
Третий закон Менделя справедлив только для тех случаев, когда гены анализируемых признаков находятся в разных парах гомологичных хромосом.
Цитологические основы третьего закона Менделя
Пусть А — ген, обусловливающий развитие желтой окраски семян, а — зеленой окраски, В — гладкая форма семени, b — морщинистая. Скрещиваются гибриды первого поколения, имеющие генотип АаВb. При образовании гамет из каждой пары аллельных генов в гамету попадает только один, при этом в результате случайного расхождения хромосом в первом делении мейоза ген А может попасть в одну гамету с геном В или с геном b, а ген а — с геном В или с геном b. Таким образом, каждый организм образует четыре сорта гамет в одинаковом количестве (по 25%): АВ, Ab, aB, ab. Во время оплодотворения каждый из четырех типов сперматозоидов может оплодотворить любую из четырех типов яйцеклеток. В результате оплодотворения возможно появление девяти генотипических классов, которые дадут четыре фенотипических класса.
Сцепленное наследование
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.
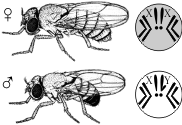
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1.
Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
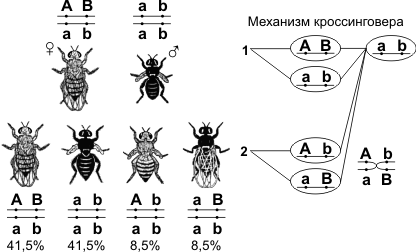
1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb.
Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот.
Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
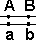 | Образуются гаметы: | 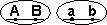 |
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
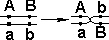 | Образуются гаметы: | 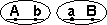 |
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза).
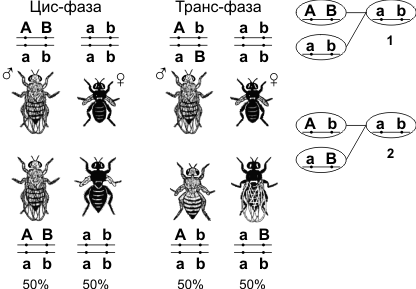
1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы).
Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности:
· гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален;
· каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
· гены расположены в хромосомах в определенной линейной последовательности;
· гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления;
· число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
· сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом;
· частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера;
· каждый вид имеет характерный только для него набор хромосом — кариотип.
Генетика человека (антропогенетика)
1. Методы изучения наследственности человека: генеалогические, близнецовые, цитогенетические, биохимические и популяционные
2. Генетические заболевания и наследственные болезни. Значение медико-генетических консультаций и пренатальной диагностики. Возможности генетической коррекции заболеваний
Генетика человека – это особый раздел генетики, который изучает особенности наследования признаков у человека, наследственные заболевания (медицинская генетика), генетическую структуру популяций человека. Генетика человека является теоретической основой современной медицины и современного здравоохранения.
В настоящее время твердо установлено, что в живом мире законы генетики носят всеобщий характер, действительны они и для человека.
Однако, поскольку человек – это не только биологическое, но и социальное существо, генетика человека отличается от генетики большинства организмов рядом особенностей:
– для изучения наследования человека неприменим гибридологический анализ (метод скрещиваний); поэтому для генетического анализа используются специфические методы: генеалогический (метод анализа родословных), близнецовый, а также цитогенетические, биохимические, популяционные и некоторые другие методы;
– для человека характерны социальные признаки, которые не встречаются у других организмов, например, темперамент, сложные коммуникационные системы, основанные на речи, а также математические, изобразительные, музыкальные и иные способности;
– благодаря общественной поддержке возможно выживание и существование людей с явными отклонениями от нормы (в дикой природе такие организмы оказываются нежизнеспособными).
Генетика человека изучает особенности наследования признаков у человека, наследственные заболевания (медицинская генетика), генетическую структуру популяций человека. Генетика человека является теоретической основой современной медицины и современного здравоохранения. Известно несколько тысяч собственно генетических заболеваний, которые почти на 100% зависят от генотипа особи. К наиболее страшным из них относятся: кислотный фиброз поджелудочной железы, фенилкетонурия, галактоземия, различные формы кретинизма, гемоглобинопатии, а также синдромы Дауна, Тернера, Кляйнфельтера. Кроме того, существуют заболевания, которые зависят и от генотипа, и от среды: ишемическая болезнь, сахарный диабет, ревматоидные заболевания, язвенные болезни желудка и двенадцатиперстной кишки, многие онкологические заболевания, шизофрения и другие заболевания психики.
Задачи медицинской генетики заключаются в своевременном выявлении носителей этих заболеваний среди родителей, выявлении больных детей и выработке рекомендаций по их лечению. Большую роль в профилактике генетически обусловленных заболеваний играют генетико-медицинские консультации и пренатальная диагностика (то есть выявление заболеваний на ранних стадиях развития организма).
Существуют специальные разделы прикладной генетики человека (экологическая генетика, фармакогенетика, генетическая токсикология), изучающие генетические основы здравоохранения. При разработке лекарственных препаратов, при изучении реакции организма на воздействие неблагоприятных факторов необходимо учитывать как индивидуальные особенности людей, так и особенности человеческих популяций.
Приведем примеры наследования некоторых морфофизиологических признаков.
Доминантные и рецессивные признаки у человека (для некоторых признаков указаны контролирующие их гены)
| Доминантные | Рецессивные |
| Нормальная пигментация кожи, глаз, волос | Альбинизм |
| Близорукость | Нормальное зрение |
| Нормальное зрение | Ночная слепота |
| Цветовое зрение | Дальтонизм |
| Катаракта | Отсутствие катаракты |
| Косоглазие | Отсутствие косоглазия |
| Толстые губы | Тонкие губы |
| Полидактилия (добавочные пальцы) | Нормальное число пальцев |
| Брахидактилия (короткие пальцы) | Нормальная длина пальцев |
| Веснушки | Отсутствие веснушек |
| Нормальный слух | Врожденная глухота |
| Карликовость | Нормальный рост |
| Нормальное усвоение глюкозы | Сахарный диабет |
| Нормальная свертываемость крови | Гемофилия |
| Круглая форма лица (R–) | Квадратная форма лица (rr) |
| Круглый подбородок (K–) | Квадратный подбородок (kk) |
| Ямочка на подбородке (А–) | Отсутствие ямочки (аа) |
| Ямочки на щеках (D–) | Отсутствие ямочек (dd) |
| Густые брови (B–) | Тонкие брови (bb) |
| Брови не соединяются (N–) | Брови соединяются (nn) |
| Длинные ресницы (L–) | Короткие ресницы (ll) |
| Круглый нос (G–) | Заостренный нос (gg) |
| Круглые ноздри (Q–) | Узкие ноздри (qq) |
| Свободная мочка уха (S–) | Сросшаяся мочка уха (ss) |
Неполное доминирование (указаны гены, контролирующие признак)
| Признаки | Варианты | ||
| Расстояние между глазами – Т | Большое | Среднее | Малое |
| Размер глаз – Е | Большие | Средние | Маленькие |
| Размеры рта – М | Большой | Средний | Маленький |
| Тип волос – С | Курчавые | Вьющиеся | Прямые |
| Цвет бровей – Н | Очень темные | Темные | Светлые |
| Размер носа – F | Большой | Средний | Маленький |
Наследование цвета волос (контролируется четырьмя генами, наследуется полимерно)
| Количество доминантных аллелей | Цвет волос |
| Черные | |
| Темно-коричневые | |
| Темно-каштановые | |
| Каштановые | |
| Русые | |
| Светло-русые | |
| Блондин | |
| Очень светлый блондин | |
| Белые |
Примечание. Рыжий цвет волос контролируется геном D; это признак проявляется, если доминантных генов меньше 6: DD – ярко-рыжие, Dd – светло-рыжие, dd – не-рыжие
Методы изучения наследственности человека: генеалогические, близнецовые, цитогенетические, биохимические и популяционные
Генеалогические методы (методы анализа родословных)
Родословная – это схема, отражающая связи между членами семьи. Анализируя родословные, изучают какой-либо нормальный или (чаще) патологический признак в поколениях людей, находящихся в родственных связях.
Генеалогические методы используются для определения наследственного или ненаследственного характера признака, доминантности или рецессивности, картирования хромосом, сцепления с полом, для изучения мутационного процесса. Как правило, генеалогический метод составляет основу для заключений при медико-генетическом консультировании.
При составлении родословных применяют стандартные обозначения. Персона (индивидуум), с которого начинается исследование, называется пробандом (если родословная составляется таким образом, что от пробанда спускаются к его потомству, то ее называют генеалогическим древом). Потомок брачной пары называется сиблингом, родные братья и сестры – сибсами, кузены – двоюродными сибсами и т.д. Потомки, у которых имеется общая мать (но разные отцы), называются единоутробными, а потомки, у которых имеется общий отец (но разные матери) – единокровными; если же в семье имеются дети от разных браков, причем, у них нет общих предков (например, ребенок от первого брака матери и ребенок от первого брака отца), то их называют сводными.
Каждый член родословной имеет свой шифр, состоящий из римской цифры и арабской, обозначающих соответственно номер поколения и номер индивидуума при нумерации поколений последовательно слева направо. При родословной должна быть легенда, т. е. пояснение к принятым обозначениям. Фрагменты, родословных, иллюстрирующих наследование доминантных и рецессивных признаков, а также редких признаков приведены ниже (рис. 2, 3).
При близкородственных браках высока вероятность К обнаружения у супругов одного и того же неблагоприятного аллеля или хромосомной аберрации (рис. 4):
Приведем значения К [X–Y] для некоторых пар родственников при моногамии:
К [родители–потомки]=К [сибсы]=1/2;
К [дед–внук]=К [дядя–племянник]=1/4;
К [двоюродные сибсы]= К [прадед–правнук]=1/8;
К [троюродные сибсы]=1/32;
К [четвероюродные сибсы]=1/128. Обычно столь дальние родственники в составе одной семьи не рассматриваются.
На основании генеалогического анализа дается заключение о наследственной обусловленности признака. Например, детально прослежено наследование гемофилии А среди потомков английской королевы Виктории. Генеалогический анализ позволил установить, что гемофилия А – это рецессивное заболевание, сцепленное с полом.
Близнецовый метод
Близнецы – это два и более ребенка, зачатые и рожденные одной матерью почти одновременно. Термин «близнецы» используется по отношению к человеку и тем млекопитающим, у которых в норме рождается один ребенок (детеныш). Различают однояйцевых и разнояйцевых близнецов.
Однояйцевые (монозиготные, идентичные) близнецы возникают на самых ранних стадиях дробления зиготы, когда два или четыре бластомера сохраняют способность при обособлении развиться в полноценный организм. Поскольку зигота делится митозом, генотипы однояйцевых близнецов, по крайней мере, исходно, совершенно идентичны. Однояйцевые близнецы всегда одного пола, в период внутриутробного развития у них одна плацента.
Разнояйцевые (дизиготные, неидентичные) близнецы возникают иначе – при оплодотворении двух или нескольких одновременно созревших яйцеклеток. Таким образом, они имеют около 50% общих генов. Другими словами, они подобны обычным братьям и сестрам по своей генетической конституции и могут быть как однополыми, так и разнополыми.
Таким образом, сходство между однояйцевыми близнецами определяется и одинаковыми генотипами, и одинаковыми условиями внутриутробного развития. Сходство между разнояйцевыми близнецами определяется только одинаковыми условиями внутриутробного развития.
Частота рождения близнецов в относительных цифрах невелика и составляет около 1%, из них 1/3 приходится на монозиготных близнецов. Однако в пересчете на общую численность населения Земли в мире проживает свыше 30 млн. разнояйцевых и 15 млн. однояйцевых близнецов.
Для исследований на близнецах очень важно установить достоверность зиготности. Наиболее точно зиготность устанавливают с помощью реципрокной трансплантации небольших участков кожи. У дизиготных близнецов трансплантаты всегда отторгаются, тогда как у монозиготных близнецов пересаженные кусочки кожи успешно приживаются. Так же успешно и длительно функционируют трансплантированные почки, пересаженные от одного из монозиготных близнецов другому
При сравнении однояйцевых и разнояйцевых близнецов, воспитанных в одной и той же среде, можно сделать заключение о роли генов в развитии признаков. Условия послеутробного развития для каждого из близнецов могут оказаться разными. Например, монозиготные близнецы были разлучены через несколько дней после рождения и воспитывались в разных условиях. Сравнение их через 20 лет по многим внешним признакам (рост, объем головы, число бороздок на отпечатках пальцев и т. д.) выявило лишь незначительные различия. В то же время, среда оказывает воздействие на ряд нормальных и патологических признаков.
Близнецовый метод позволяет делать обоснованные заключения о наследуемости признаков: роли наследственности, среды и случайных факторов в определении тех или иных признаков человека,
Наследуемость – это вклад генетических факторов в формирование признака, выраженный в долях единицы или процентах.
Для вычисления наследуемости признаков сравнивают степень сходства или различия по ряду признаков у близнецов разного типа.
Рассмотрим некоторые примеры, иллюстрирующие сходство (конкордантность) и различие (дискордантность) многих признаков (см. табл.).
Степень различия (дискордантность) по ряду нейтральных признаков у близнецов
| Признаки, контролируемые небольшим числом генов | Частота (вероятность) появления различий, % | Наследуемость, % | |
| однояйцевые | разнояйцевые | ||
| Цвет глаз | 0,5 | ||
| Форма ушей | 2,0 | ||
| Цвет волос | 3,0 | ||
| Папиллярные линии | 8,0 | ||
| среднее | < 1 % | ≈ 55 % | 95 % |
| Биохимические признаки | 0,0 | от 0 до 100 | 100 % |
| Цвет кожи | 0,0 | ||
| Форма волос | 0,0 | ||
| Форма бровей | 0,0 | ||
| Форма носа | 0,0 | ||
| Форма губ | 0,0 |
Степень сходства (конкордантность) по ряду заболеваний у близнецов
| Признаки, контролируемые большим числом генов и зависящие от негенетических факторов | Частота (вероятность) появления сходства, % | Наследуемость, % | |
| однояйцевые | разнояйцевые | ||
| Умственная отсталость | |||
| Шизофрения | |||
| Сахарный диабет | |||
| Эпилепсия | |||
| среднее | ≈ 70 % | ≈ 20 % | ≈ 65 % |
| Преступность (?) | 56 % |
Обращает на себя внимание высокая степень сходства однояйцевых близнецов по таким тяжелым заболеваниям, как шизофрения, эпилепсия, сахарный диабет.
Кроме морфологических признаков, а также тембра голоса, походки, мимики, жестикуляции и т. д. изучают антигенную структуру клеток крови, белки сыворотки, способность ощущать вкус некоторых веществ.
Особый интерес представляет наследование социально значимых признаков: агрессивности, альтруизма, творческих, исследовательских, организаторских способностей. Считается, что социально значимые признаки примерно на 80 % обусловлены генотипом.
Цитогенетические (кариотипические, кариотипические) методы
Цитогенетические методы используются, в первую очередь, при изучении кариотипов отдельных индивидов. Кариотип человека довольно хорошо изучен (рис. 6). Применение дифференциальной окраски позволяет точно идентифицировать все хромосомы. Общее число хромосом в гаплоидном наборе равно 23. Из них 22 хромосомы одинаковы и у мужчин, и у женщин; они называются аутосомы. В диплоидном наборе (2n=46) каждая аутосома представлена двумя гомологами. Двадцать третья хромосома является половой хромосомой, она может быть представлена или X или Y–хромосомой. Половые хромосомы у женщин представлены двумя X–хромосомами, а у мужчин одной X–хромосомой и одной Y–хромосомой.
Изменение кариотипа, как правило, связано с развитием генетических заболеваний (см. ниже).
Благодаря культивированию клеток человека in vitro можно быстро получить достаточно большой материал для приготовления препаратов. Для кариотипирования обычно используют кратковременную культуру лейкоцитов периферической крови.
Цитогенетические методы используются и для описания интерфазных клеток. Например, по наличию или отсутствию полового хроматина (телец Барра, представляющих собой инактивированные X-хромосомы) можно не только определять пол индивидов, но и выявлять некоторые генетические заболевания, связанные с изменением числа X-хромосом (см. ниже).
Картирование хромосом человека.
Для картирования генов человека широко используются методы биотехнологии. В частности, методы клеточной инженерии позволяют объединять различные типы клеток. Слияние клеток, принадлежащих к разным биологическим видам, называется соматической гибридизацией. Сущность соматической гибридизации заключается в получении синтетических культур путем слияния протопластов различных видов организмов. Для слияния клеток используют различные физико-химические и биологические методы. После слияния протопластов образуются многоядерные гетерокариотические клетки. В дальнейшем при слиянии ядер образуются синкариотические клетки, содержащие в ядрах хромосомные наборы разных организмов. При делении таких клеток in vitro образуются гибридные клеточные культуры. В настоящее время получены и культивируются клеточные гибриды «человек × мышь», «человек × крыса» и многие другие.
В гибридных клетках, полученных из разных штаммов разных видов, один из родительских геномов постепенно теряет хромосомы. Эти процессы интенсивно протекают, например, в клеточных гибридах между мышью и человеком. Если при этом следить за каким-либо биохимическим маркером (например, определенным ферментом человека) и одновременно проводить цитогенетический контроль, то, в конце концов, можно связать исчезновение хромосомы одновременно с биохимическим признаком. Это означает, что ген, кодирующий этот признак, локализован в данной хромосоме.
Дополнительная информация о локализации генов может быть получена при анализе хромосомных мутаций (делеций).
Биохимические методы.
Все многообразие биохимических методов делится на две группы.
а) Методы, основанные на выявлении определенных биохимических продуктов, обусловленных действием разных аллелей. Легче всего выявлять аллели по изменению активности ферментов или по изменению какого-либо биохимического признака.
б) Методы, основанные на непосредственном выявлении измененных нуклеиновых кислот и белков с помощью гель-электрофореза в сочетании с другими методиками (блот-гибридизации, авторадиографии).
Использование биохимических методов позволяет выявить гетерозиготных носителей заболеваний. Например, у гетерозиготных носителей гена фенилкетонурии изменяется уровень фенилаланина в крови.
Методы генетики мутагенеза
Мутационный процесс у человека у человека, как и у всех других организмов, ведет к возникновению аллелей и хромосомных перестроек, отрицательно влияющих на здоровье.
Генные мутации. Около 1% новорожденных заболевают вследствие генных мутаций, из которых часть вновь возникшие. Темп мутирования различных генов в генотипе человека неодинаков. Известны гены, которые мутирует с частотой 10–4 на гамету на поколение. Однако большинство других генов мутируют с частотой, в сотни раз меньшей (10–6). Ниже приведены примеры наиболее частых генных мутаций у человека:
| Типы и названия мутаций | Частота мутаций (на 1 млн. гамет) |
| Аутосомно-доминантные | |
| Поликистоз почек | 65...120 |
| Нейрофиброматоз | 11...100 |
| Множественный полипоз толстой кишки | 10...50 |
| Аномалия лейкоцитов Пельгера | 9...27 |
| Несовершенный остеогенез | 7...13 |
| Синдром Марфана | 4...6 |
| Аутосомно-рецессивные | |
| Микроцефалия | |
| Ихтиоз (не сцепленный с полом) | |
| Рецессивные, сцепленные с полом | |
| Мышечная дистрофия Дюшена | 43...105 |
| Гемофилия А | 37...52 |
| Гемофилия В | 2...3 |
| Ихтиоз (сцепленный с полом) |
Хромосомные и геномные мутации в абсолютном большинстве возникают в половых клетках родителей. Один из 150 новорожденных несет хромосомную мутацию. Около 50% ранних абортов обусловлено хромосомными мутациями. Это связано с тем, что одна из 10 гамет человека является носителем структурных мутаций. Возраст родителей, особенно возраст матерей, играет важную роль в увеличении частоты хромосомных, а возможно, и генных мутаций.
Полиплоидия у человека встречается очень редко. Известны случая рождения триплоидов – эти новорожденные рано умирают. Тетраплоиды обнаружены среди абортированных зародышей.
В настоящее время мутационный процесс у человека характеризуется тем, что протекает на фоне повышенной концентрации мутагенных факторов, созданной производственной деятельностью самого человека. Важнейшая задача сегодняшнего дня – выявление мутагенных свойств загрязнителей, особенно новых химических веществ (лекарств, пестицидов, пищевых добавок, различных видов топлива и т.д.), и разработка методов технологии, позволяющих предотвратить возникновение опасных концентраций этих агентов. Одним из сильнейших мутагенов является радиация (ионизирующие излучения). Доказано, что не существует пороговой дозы ионизирующих излучений. Другими словами, индукция мутаций может быть достигнута при действии любых доз, а при увеличении дозы пропорционально растет число мутаций. Мутагенным действием на клетки человека обладают и некоторые вирусы, причем даже в ослабленной форме, которая используется для приготовления вакцин. Известно также, что большинство мутагенов обладают и канцерогенными свойствами, то есть они могут индуцировать развитие злокачественных опухолей (см. ниже).
В то же время существуют факторы, которые сн

