 2015-04-30
2015-04-30 3536
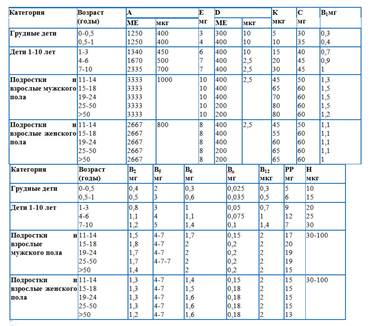
3536- Витамин А (ретинол);
- Витамин D (холекальциферол);
- Витамин Е (токоферол);
- Витамин К (филлохинон).
15. Витамин А, структура, участие в обменных процессах. Гипо- гипер- и авитаминоз А, их причины и особенности проявления. Пищевые источники, суточная потребность.
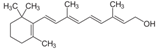 Витамин А (ретинол) - циклический, ненасыщенный, одноатомный спирт. Витамин А содержится только в животных продуктах: печени крупного рогатого скота и свиней, яичном желтке, молочных продуктах; особенно богат этим витамином рыбий жир. Суточная потребность витамина А взрослого человека составляет от 1 до 2,5 мг витамина. Биологические функции витамина А. В организме ретинол превращается в ретиналь и ретиноевую кислоту, участвующие в регуляции ряда функций (в росте и дифференцировке клеток); они также составляют фотохимическую основу акта зрения. Основные клинические проявления гиповитаминоза А. Наиболее ранний и характерный признак недостаточности витамина А у людей - нарушение сумеречного зрения (гемералопия, или "куриная" слепота). Специфично для авитаминоза А поражение глазного яблока - ксерофтальмия, т.е. развитие сухости роговой оболочки глаза как следствие закупорки слёзного канала в связи с ороговением эпителия. Это, в свою очередь, приводит к развитию конъюнктивита, отёку, изъязвлению и размягчению роговой оболочки, т.е. к кератомаляции. Ксерофтальмия и кератомаляция при отсутствии соответствующего лечения могут привести к полной потере зрения.
Витамин А (ретинол) - циклический, ненасыщенный, одноатомный спирт. Витамин А содержится только в животных продуктах: печени крупного рогатого скота и свиней, яичном желтке, молочных продуктах; особенно богат этим витамином рыбий жир. Суточная потребность витамина А взрослого человека составляет от 1 до 2,5 мг витамина. Биологические функции витамина А. В организме ретинол превращается в ретиналь и ретиноевую кислоту, участвующие в регуляции ряда функций (в росте и дифференцировке клеток); они также составляют фотохимическую основу акта зрения. Основные клинические проявления гиповитаминоза А. Наиболее ранний и характерный признак недостаточности витамина А у людей - нарушение сумеречного зрения (гемералопия, или "куриная" слепота). Специфично для авитаминоза А поражение глазного яблока - ксерофтальмия, т.е. развитие сухости роговой оболочки глаза как следствие закупорки слёзного канала в связи с ороговением эпителия. Это, в свою очередь, приводит к развитию конъюнктивита, отёку, изъязвлению и размягчению роговой оболочки, т.е. к кератомаляции. Ксерофтальмия и кератомаляция при отсутствии соответствующего лечения могут привести к полной потере зрения.
16. Витамин Д, структура, участие в обменных процессах. Активные формы витамина Д в организме. Гипер-, гипо- и авитаминоз Д. Пищевые источники, суточная потребность.Роль витамина Д как антирахитического фактора. Биохимическая сущность рахита.
 Витамины группы D (кальциферолы) - группа химически родственных соединений, относящихся к производным стеринов. Наиболее биологически активные витамины - D2 и D3. Витамин D2 (эргокалферол), производное эргостерина - растительного стероида, встречающегося в некоторых грибах, дрожжах и растительных маслах. Источники. Наибольшее количество витамина D3 содержится в продуктах животного происхождения: сливочном масле, желтке яиц, рыбьем жире. Суточная потребность для детей 12-25 мкг (500-1000 ME), для взрослого человека потребность значительно меньше. Биологическая роль. В организме человека витамин D3 гидроксилируется в положениях 25 и 1 и превращается в биологически активное соединение 1,25-дигидроксихолекальциферол. Кальцитриол выполняет гормональную функцию, участвуя в регуляции обмена Са2+ и фосфатов, стимулируя всасывание Са2+ в кишечнике и кальцификацию костной ткани, реабсорбцию Са2+и фосфатов в почках. Недостаточность. При недостатке витамина D у детей развивается заболевание "рахит", характеризуемое нарушением кальцификации растущих костей. При этом наблюдают деформацию скелета с характерными изменениями. Избыток. Поступление в организм избыточного количества витамина D3 может вызвать гипервитаминоз D. Это состояние характеризуется избыточным отложением солей кальция в тканях лёгких, почек, сердца, стенках сосудов, а также остеопорозом с частыми переломами костей.
Витамины группы D (кальциферолы) - группа химически родственных соединений, относящихся к производным стеринов. Наиболее биологически активные витамины - D2 и D3. Витамин D2 (эргокалферол), производное эргостерина - растительного стероида, встречающегося в некоторых грибах, дрожжах и растительных маслах. Источники. Наибольшее количество витамина D3 содержится в продуктах животного происхождения: сливочном масле, желтке яиц, рыбьем жире. Суточная потребность для детей 12-25 мкг (500-1000 ME), для взрослого человека потребность значительно меньше. Биологическая роль. В организме человека витамин D3 гидроксилируется в положениях 25 и 1 и превращается в биологически активное соединение 1,25-дигидроксихолекальциферол. Кальцитриол выполняет гормональную функцию, участвуя в регуляции обмена Са2+ и фосфатов, стимулируя всасывание Са2+ в кишечнике и кальцификацию костной ткани, реабсорбцию Са2+и фосфатов в почках. Недостаточность. При недостатке витамина D у детей развивается заболевание "рахит", характеризуемое нарушением кальцификации растущих костей. При этом наблюдают деформацию скелета с характерными изменениями. Избыток. Поступление в организм избыточного количества витамина D3 может вызвать гипервитаминоз D. Это состояние характеризуется избыточным отложением солей кальция в тканях лёгких, почек, сердца, стенках сосудов, а также остеопорозом с частыми переломами костей.
17. Витамин РР, структура коферментов, участие в обменных процессах. Гипо - и авитаминоз РР. Пищевые источники, суточная потребность.
 Витамин РР (никотиновая кислота, никотинамид, витамин B3) Источники. Витамин РР широко распространён в растительных продуктах, высоко его содержание в рисовых и пшеничных отрубях, дрожжах, много витамина в печени и почках крупного рогатого скота и свиней. Суточная потребность в этом витамине доставляет для взрослых 15-25 мг, для детей - 15 мг. Биологические функции. Никотиновая кислота в организме входит в состав NAD и NADP, выполняющих функции коферментов различных дегидрогеназ. Недостаточность витамина РР приводит к заболеванию "пеллагра", для которого характерны 3 основных признака: дерматит, диарея, деменция ("три Д"), Пеллагра проявляется в виде симметричного дерматита на участках кожи, доступных действию солнечных лучей, расстройств ЖКТ (диарея) и воспалительных поражений слизистых оболочек рта и языка. В далеко зашедших случаях пеллагры наблюдают расстройства ЦНС (деменция): потеря памяти, галлюцинации и бред.
Витамин РР (никотиновая кислота, никотинамид, витамин B3) Источники. Витамин РР широко распространён в растительных продуктах, высоко его содержание в рисовых и пшеничных отрубях, дрожжах, много витамина в печени и почках крупного рогатого скота и свиней. Суточная потребность в этом витамине доставляет для взрослых 15-25 мг, для детей - 15 мг. Биологические функции. Никотиновая кислота в организме входит в состав NAD и NADP, выполняющих функции коферментов различных дегидрогеназ. Недостаточность витамина РР приводит к заболеванию "пеллагра", для которого характерны 3 основных признака: дерматит, диарея, деменция ("три Д"), Пеллагра проявляется в виде симметричного дерматита на участках кожи, доступных действию солнечных лучей, расстройств ЖКТ (диарея) и воспалительных поражений слизистых оболочек рта и языка. В далеко зашедших случаях пеллагры наблюдают расстройства ЦНС (деменция): потеря памяти, галлюцинации и бред.
18. Витамин В2, структура коферментов, участие в обменных процессах. Гиповитаминоз и авитаминоз В2. Суточная потребность, пищевые источники.
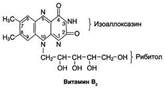 Витамин В2 (рибофлавин). В основе структуры витамина В2 лежит структура изоаллоксазина, соединённого со спиртом рибитолом. Главные источники витамина В2 - печень, почки, яйца, молоко, дрожжи. Витамин содержится также в шпинате, пшенице, ржи. Частично человек получает витамин В2 как продукт жизнедеятельности кишечной микрофлоры. Суточная потребность в витамине В2 взрослого человека составляет 1,8-2,6 мг. Биологические функции. В слизистой оболочке кишечника после всасывания витамина происходит образование коферментов FMN и FAD. Клинические проявления недостаточности рибофлавина выражаются в остановке роста у молодых организмов. Часто развиваются воспалительные процессы на слизистой оболочке ротовой полости, появляются длительно незаживающие трещины в углах рта, дерматит носогубной складки. Типично воспаление глаз: конъюнктивиты, васкуляризация роговицы, катаракта. Кроме того, при авитаминозе В2 развиваются общая мышечная слабость и слабость сердечной мышцы.
Витамин В2 (рибофлавин). В основе структуры витамина В2 лежит структура изоаллоксазина, соединённого со спиртом рибитолом. Главные источники витамина В2 - печень, почки, яйца, молоко, дрожжи. Витамин содержится также в шпинате, пшенице, ржи. Частично человек получает витамин В2 как продукт жизнедеятельности кишечной микрофлоры. Суточная потребность в витамине В2 взрослого человека составляет 1,8-2,6 мг. Биологические функции. В слизистой оболочке кишечника после всасывания витамина происходит образование коферментов FMN и FAD. Клинические проявления недостаточности рибофлавина выражаются в остановке роста у молодых организмов. Часто развиваются воспалительные процессы на слизистой оболочке ротовой полости, появляются длительно незаживающие трещины в углах рта, дерматит носогубной складки. Типично воспаление глаз: конъюнктивиты, васкуляризация роговицы, катаракта. Кроме того, при авитаминозе В2 развиваются общая мышечная слабость и слабость сердечной мышцы.
19. Витамин С, структура, участие в обменных процессах. Гипо- и авитаминоз С. Пищевые источники, суточная потребность.
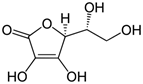 Витамин С (аскорбиновая кислота). Источники витамина С - свежие фрукты, овощи, зелень. Суточная потребность человека в витамине С составляет 50-75 мг. Биологические функции. Образование коллагена, серотонина из триптофана, образование катехоламинов, синтез кортикостероидов. Аскорбиновая кислота также участвует в превращении холестерина в желчные кислоты. Витамин С необходим для детоксикации в гепатоцитах при участии цитохрома P450. Витамин С сам нейтрализует супероксид-анион радикал до перекиси водорода. Клинические проявления недостаточности витамина С. Недостаточность аскорбиновой кислоты приводит к заболеванию, называемому цингой (скорбут). Цинга, возникающая у человека при недостаточном содержании в пищевом рационе свежих фруктов и овощей, описана более 300 лет назад, со времени проведения длительных морских плаваний и северных экспедиций. Это заболевание связано с недостатком в пище витамина С. Болеют цингой только человек, приматы и морские свинки. Главные проявления авитаминоза обусловлены в основном нарушением образования коллагена в соединительной ткани. Вследствие этого наблюдают разрыхление дёсен, расшатывание зубов, нарушение целостности капилляров (сопровождающееся подкожными кровоизлияниями). Возникают отёки, боль в суставах, анемия.
Витамин С (аскорбиновая кислота). Источники витамина С - свежие фрукты, овощи, зелень. Суточная потребность человека в витамине С составляет 50-75 мг. Биологические функции. Образование коллагена, серотонина из триптофана, образование катехоламинов, синтез кортикостероидов. Аскорбиновая кислота также участвует в превращении холестерина в желчные кислоты. Витамин С необходим для детоксикации в гепатоцитах при участии цитохрома P450. Витамин С сам нейтрализует супероксид-анион радикал до перекиси водорода. Клинические проявления недостаточности витамина С. Недостаточность аскорбиновой кислоты приводит к заболеванию, называемому цингой (скорбут). Цинга, возникающая у человека при недостаточном содержании в пищевом рационе свежих фруктов и овощей, описана более 300 лет назад, со времени проведения длительных морских плаваний и северных экспедиций. Это заболевание связано с недостатком в пище витамина С. Болеют цингой только человек, приматы и морские свинки. Главные проявления авитаминоза обусловлены в основном нарушением образования коллагена в соединительной ткани. Вследствие этого наблюдают разрыхление дёсен, расшатывание зубов, нарушение целостности капилляров (сопровождающееся подкожными кровоизлияниями). Возникают отёки, боль в суставах, анемия.
20. Витамин В1, структура кофермента, участие в обменных процессах. Гиповитаминоз и авитаминоз В1. Пищевые источники, суточная потребность.
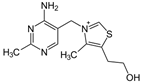 Витамин B1 (тиамин). Структура витамина включает пиримидиновое и тиазоловое кольца, соединённые метановым мостиком. Источники. Он широко распространён в продуктах растительного происхождения (оболочка семян хлебных злаков и риса, горох, фасоль, соя и др.). Суточная потребность взрослого человека в среднем составляет 2-3 мг витамина В1. Биологическая роль витамина В, определяется тем, что в виде ТДФ он входит в состав как минимум трёх ферментов и ферментных комплексов: в составе пируват - и оскетоглутаратдегидрогеназных комплексов он участвует в окислительном декарбоксилировании пирувата и оскетоглутарата; в составе транскетолазы ТДФ участвует в пентозофосфатном пути превращения углеводов. Основной, наиболее характерный и специфический признак недостаточности витамина В1 - полиневрит, в основе которого лежат дегенеративные изменения нервов. Вначале развивается болезненность вдоль нервных стволов, затем - потеря кожной чувствительности и наступает паралич (бери-бери). Второй важнейший признак заболевания - нарушение сердечной деятельности, что выражается в нарушении сердечного ритма, увеличении размеров сердца и в появлении болей в области сердца. К характерным признакам заболевания, связанного с недостаточностью витамина В1относят также нарушения секреторной и моторной функций ЖКТ; наблюдают снижение кислотности желудочного сока, потерю аппетита, атонию кишечника.
Витамин B1 (тиамин). Структура витамина включает пиримидиновое и тиазоловое кольца, соединённые метановым мостиком. Источники. Он широко распространён в продуктах растительного происхождения (оболочка семян хлебных злаков и риса, горох, фасоль, соя и др.). Суточная потребность взрослого человека в среднем составляет 2-3 мг витамина В1. Биологическая роль витамина В, определяется тем, что в виде ТДФ он входит в состав как минимум трёх ферментов и ферментных комплексов: в составе пируват - и оскетоглутаратдегидрогеназных комплексов он участвует в окислительном декарбоксилировании пирувата и оскетоглутарата; в составе транскетолазы ТДФ участвует в пентозофосфатном пути превращения углеводов. Основной, наиболее характерный и специфический признак недостаточности витамина В1 - полиневрит, в основе которого лежат дегенеративные изменения нервов. Вначале развивается болезненность вдоль нервных стволов, затем - потеря кожной чувствительности и наступает паралич (бери-бери). Второй важнейший признак заболевания - нарушение сердечной деятельности, что выражается в нарушении сердечного ритма, увеличении размеров сердца и в появлении болей в области сердца. К характерным признакам заболевания, связанного с недостаточностью витамина В1относят также нарушения секреторной и моторной функций ЖКТ; наблюдают снижение кислотности желудочного сока, потерю аппетита, атонию кишечника.
21. Витамин В6, структура кофермента, участие в обменных процессах. Гиповитаминоз и авитаминоз В6. Пищевые источники, суточная потребность.
ВитаминВ6 (пиридоксин, пиридоксаль, пиридоксамин) В основе структуры витамина В6 лежит пиридиновое кольцо. Известны 3 формы витамина В6, отличающиеся строением замещающей группы у атома углерода в п-положении к атому азота. Все они характеризуются одинаковой биологической активностью. 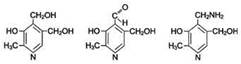 Источники витамина В6 для человека - такие продукты питания, как яйца, печень, молоко, зеленый перец, морковь, пшеница, дрожжи. Некоторое количество витамина синтезируется кишечной флорой. Суточная потребностьсоставляет 2-3 мг. Биологические функции.Все формы витамина В6 используются в организме для синтеза коферментов: пиридоксальфосфата и пиридоксаминфосфата. Коферменты образуются путём фосфорилирования по гидроксиметильной группе в пятом положении пиримидинового кольца при участии фермента пиридоксалькиназы и АТФ как источника фосфата. Клинические проявления недостаточности витамина. Авитаминоз В6 у детей проявляется повышенной возбудимостью ЦНС, периодическими судорогами, что связано, возможно, с недостаточным образованием тормозного медиатора ГАМК, специфическими дерматитами. У взрослых признаки гиповитаминоза В6 наблюдают при длительном лечении туберкулёза изониазидом (антагонист витамина В6). При этом возникают поражения нервной системы (полиневриты), дерматиты.
Источники витамина В6 для человека - такие продукты питания, как яйца, печень, молоко, зеленый перец, морковь, пшеница, дрожжи. Некоторое количество витамина синтезируется кишечной флорой. Суточная потребностьсоставляет 2-3 мг. Биологические функции.Все формы витамина В6 используются в организме для синтеза коферментов: пиридоксальфосфата и пиридоксаминфосфата. Коферменты образуются путём фосфорилирования по гидроксиметильной группе в пятом положении пиримидинового кольца при участии фермента пиридоксалькиназы и АТФ как источника фосфата. Клинические проявления недостаточности витамина. Авитаминоз В6 у детей проявляется повышенной возбудимостью ЦНС, периодическими судорогами, что связано, возможно, с недостаточным образованием тормозного медиатора ГАМК, специфическими дерматитами. У взрослых признаки гиповитаминоза В6 наблюдают при длительном лечении туберкулёза изониазидом (антагонист витамина В6). При этом возникают поражения нервной системы (полиневриты), дерматиты.
22. Пантотеновая кислота и биотин, их участие в обменных процессах. Гиповитаминозы и авитаминозы В3 и Н. Суточная потребность, источник этих витаминов.
Пантотеновая кислота (лекарственная форма — пантотенат кальция, витамин B5, неверное обозначение — витамин B3). Пантотеновая кислота требуется для обмена жиров, углеводов, аминокислот, синтеза жизненно важных жирных кислот, холестерина, гистамина, ацетилхолина, гемоглобина. Пантотеновая кислота чувствительна к нагреванию, при термической обработке теряется почти 50 % витамина. Суточная потребность человека в пантотеновой кислоте (5—10 мг) удовлетворяется при нормальном смешанном питании, так как пантотеновая кислота содержится в очень многих продуктах животного и растительного происхождения (дрожжи, икра рыб, яичный желток, зелёные части растений, молоко, морковь, капуста и т. д.). Симптомы гиповитаминоза: 1.усталость 2.депрессия 3.расстройство сна 4.повышенная утомляемость 5.головные боли 6. Тошнота 7. мышечные боли
23. Фолиевая кислота и кобаламин, их участие в обменных процессах, авитаминозы. Пищевые источники, суточная потребность.
Фолиевая кислота (витамин Вc, витамин B9). Источники. Значительное количество этого витамина содержится в дрожжах, а также в печени, почках, мясе и других продуктах животного происхождения. Суточная потребность в фолиевой кислоте колеблется от 50 до 200 мкг; однако вследствие плохой всасываемости этого витамина рекомендуемая суточная доза - 400 мкг. Биологическая роль фолиевой кислоты определяется тем, что она служит субстратом для синтеза коферментов, участвующих в реакциях переноса одноуглеродных радикалов различной степени окисленности: метальных, оксиметильных, формильных и других. Эти коферменты участвуют в синтезе различных веществ: пуриновых нуклеотидов, превращении сУМФ в сПГМФ, в обмене глицина и серина. Наиболее характерные признаки авитаминоза фолиевой кислоты - нарушение кроветворения и связанные с этим различные формы малокровия (макроцитарная анемия), лейкопения и задержка роста. При гиповитаминозе фолиевой кислоты наблюдают нарушения регенерации эпителия, особенно в ЖКТ. Витамин В12 (кобаламин). Источники. Это единственный витамин, синтезируемый почти исключительно микроорганизмами: бактериями, актиномицетами и сине-зелёными водорослями. Из животных тканей наиболее богаты витамином В12 печень и почки. Суточная потребн ость в витамине В12 крайне мала и составляет всего 1-2 мкг. Биологическая роль. Витамин В12 служит источником образования двух коферментов: метилкобаламина в цитоплазме и дезоксиаденозилкобаламина в митохондриях. Основной признак авитаминоза В12 - макроцитарная (мегалобластная) анемия. Для этого заболевания характерны увеличение размеров эритроцитов, снижение количества эритроцитов в кровотоке, снижение концентрации гемоглобина в крови. Нарушение кроветворения связано в первую очередь с нарушением обмена нуклеиновых кислот, в частности синтеза ДНК в быстроделящихся клетках кроветворной системы.
24. Витамин Е и К, участие их в обменных процессах. Гиповитаминозы и авитаминозы этих витаминов. Пищевые источники, суточная потребность.
Витамины группы Е (токоферолы) найдены в природных источниках. Все они - метальные производные исходного соединения токола, по строению очень близки и обозначаются буквами греческого алфавита. Наибольшую биологическую активность проявляет α-токоферол. Источники витамина Е для человека - растительные масла, салат, капуста, семена злаков, сливочное масло, яичный желток. Суточная потребность взрослого человека в витамине примерно 5 мг. Биологическая роль. По механизму действия токоферол является биологическим антиоксидантом. Он ингибирует свободнорадикальные реакции. Клинические проявления недостаточностивитамина Е. Дефицит витамина Е проявляется развитием гемолитической анемии.
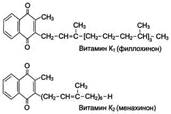 Витамины К (нафтохиноны) существует в нескольких формах в растениях как филлохинон (К1), клетках кишечной флоры как менахинон (К2). Источники витамина К - растительные (капуста, шпинат, корнеплоды и фрукты) и животные (печень) продукты. Суточная потребность в витамине взрослого составляет 1-2 мг. Биологическая функция витамина К связана с его участием в процессе свёртывания крови. Он участвует в активации факторов свёртывания крови. Витамин К участвует в реакциях карбоксилирования в качестве кофермента. Основное проявление авитаминоза К - сильное кровотечение, часто приводящее к шоку и гибели организма.
Витамины К (нафтохиноны) существует в нескольких формах в растениях как филлохинон (К1), клетках кишечной флоры как менахинон (К2). Источники витамина К - растительные (капуста, шпинат, корнеплоды и фрукты) и животные (печень) продукты. Суточная потребность в витамине взрослого составляет 1-2 мг. Биологическая функция витамина К связана с его участием в процессе свёртывания крови. Он участвует в активации факторов свёртывания крови. Витамин К участвует в реакциях карбоксилирования в качестве кофермента. Основное проявление авитаминоза К - сильное кровотечение, часто приводящее к шоку и гибели организма.
25. Возрастные потребности в отдельных витаминах у ребенка. Особенности проявления гипо- и авитаминозов в раннем детском возрасте.
Гиповитаминоз - болезненное состояние, возникающее при нарушении соответствия между расходованием витаминов и поступлением их в организм; то же, что витаминная недостаточность.
Гиповитаминоз развивается при недостаточном поступлении витаминов. Гиповитаминоз развивается незаметно: появляется раздражительность, повышенная утомляемость, снижается внимание, ухудшается аппетит, нарушается сон. Систематический длительный недостаток витаминов в пище снижает работоспособность, сказывается на состоянии отдельных органов и тканей (кожа, слизистые, мышцы, костная ткань) и важнейших функциях организма, таких как рост, интеллектуальные и физические возможности, продолжение рода, защитные силы организма. Основные причины гиповитаминозов:
- Недостаток витаминов в пище;
- Нарушение всасывания в ЖКТ;
- Врождённые дефекты ферментов, участвующих в превращениях витаминов;
- Действие структурных аналогов витаминов (антивитамины).
 Авитаминоз — заболевание, являющееся следствием длительного неполноценного питания, в котором отсутствуют какие-либо витамины.
Авитаминоз — заболевание, являющееся следствием длительного неполноценного питания, в котором отсутствуют какие-либо витамины.
Признаки авитаминоза:
§ шелушение кожного покрова.
§ ломкость ногтей.
§ выпадение волос.
Причины, вызывающие авитаминоз:
§ Нарушение поступления витаминов с пищей при неправильном питании, недостаточном или некачественном питании.
§ Нарушение процессов пищеварения или нарушение работы органов, связанных непосредственно с пищеварением.
§ Поступление в организм антивитаминов, например лекарственных препаратов синкумар, дикумарол, применяющихся при лечении повышенной свертываемости крови.
§ Особенности детского обмена веществ
§ Особенности обмена веществ у пожилых людей
Некоторые заболевания:
§ цинга — при отсутствии витамина С
§ куриная слепота - Витамин А
§ бери-бери — Витамин B1
§ рахит — Витамин D
26. Гормоны – регуляторы обменных процессов в организме. Общие биологические признаки гормонов. Классификация по их химической структуре. механизм действия гормонов. Гормоны проникающего действия и непроникающего. Циклические посредники, их роль в регуляции гормонального эффекта. Аденилатциклазная система. Роль кальция, инозилтрифосфата в передаче гормонального сигнала.
Гормоны – вещества органической природы, вырабатывающиеся в специализированных клетках желез внутренней секреции, поступающие в кровь и оказывающие регулирующее влияние на обмен веществ и физиологические функции.
В это определение необходимо внести соответствующие коррективы в связи с обнаружением типичных гормонов млекопитающих у одноклеточных (например, инсулин у микроорганизмов) или возможностью синтеза гормонов соматическими клетками в культуре ткани (например, лимфоцитами под действием факторов роста). Гормоны классифицируют в зависимости от места их природного синтеза, в соответствии с которым различают гормоны гипоталамуса, гипофиза, щитовидной железы, надпочечников, поджелудочной железы, половых желез, зобной железы и др. Современная классификация гормонов, основанной на их химической природе. В соответствии с этой классификацией различают три группы истинных гормонов:
1) пептидные и белковые гормоны,
2) гормоны – производные аминокислот
3) гормоны стероидной природы.
Четвертую группу составляют эйкозаноиды – гормоноподобные вещества, оказывающие местное действие.
Пептидные и белковые гормоны включают от 3 до 250 и более аминокислотных остатков. Это гормоны гипоталамуса и гипофиза (тиролиберин, соматолиберин, соматостатин, гормон роста, кортикотропин, тиреотропин), а также гормоны поджелудочной железы (инсулин, глюкагон). Гормоны – производные амино кислот в основном представлены производными аминокислоты тирозина. Это низкомолекулярные соединения адреналин и норадреналин, синтезирующиеся в мозговом веществе надпочечников, и гормоны щитовидной железы (тироксин и его производные). Гормоны 1-й и 2-й групп хорошо растворимы в воде. Гормоны стероидной природы представлены жирорастворимыми гормонами коркового вещества надпочечников (кортикостероиды), половыми гормонами (эстрогены и андрогены), а также гормональной формой витамина D. Эйкозаноиды, являющиеся производными полиненасыщенной жирной кислоты (арахидоновой), представлены тремя подклассами соединений: простагландины, тромбоксаны и лейкотриены. Эти нерастворимые в воде и нестабильные соединения оказывают свое действие на клетки, находящиеся вблизи их места синтеза. 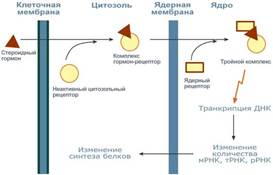 Гормоны первого типа связываются поверхностными рецепторами, расположенными на плазматической мембране. Различают три вида взаимодействия гормонов с плазматической мембраной. При взаимодействии первого вида гормонрецепторный комплекс, находящийся на поверхности клеток, вызывает образование так называемого второго посредника — циклического аденозин-3,5-монофосфата (цАМФ), и последующие действия гормона опосредуются цАМФ. Этот механизм характерен для некоторых белковых гормонов и биогенных аминов. При взаимодействии второго вида рецептор клеточной поверхности индуцирует продукцию или высвобождение иных вторых посредников, например кальция. Этот механизм характерен для некоторых нейротрансмиттеров и РТГ. При взаимодействии третьего вида комплекс поверхностный рецептор — гормон интернализуется внутрь клетки. К последней категории гормонов относится инсулин. Некоторые гормоны, такие как стероидные гормоны и гормоны щитовидной железы, по свойствам являются гидрофобными. В плазме крови для их транспортировки используются специальные белки-транспортеры. В комплексе с этими белками они не способны взаимодействовать с мембранными рецепторами, но способны отрываться от них и диффундировать через клеточную мембрану внутрь клетки. После перехода в цитозоль гормоны немедленно подхватываются другими белками, которые уже являются рецепторами. Комплекс гормон-рецептор в некоторых случаях дополнительно модифицируется и активируется. Далее он проникает в ядро, где может связываться с ядерным рецептором. В результате гормон приобретает сродство к ДНК. Связываясь с гормон-чувствительным элементом в ДНК, гормон влияет на транскрипцию определенных генов и изменяет концентрацию РНК в клетке и, соответственно, количество определенных белков в клетке.
Гормоны первого типа связываются поверхностными рецепторами, расположенными на плазматической мембране. Различают три вида взаимодействия гормонов с плазматической мембраной. При взаимодействии первого вида гормонрецепторный комплекс, находящийся на поверхности клеток, вызывает образование так называемого второго посредника — циклического аденозин-3,5-монофосфата (цАМФ), и последующие действия гормона опосредуются цАМФ. Этот механизм характерен для некоторых белковых гормонов и биогенных аминов. При взаимодействии второго вида рецептор клеточной поверхности индуцирует продукцию или высвобождение иных вторых посредников, например кальция. Этот механизм характерен для некоторых нейротрансмиттеров и РТГ. При взаимодействии третьего вида комплекс поверхностный рецептор — гормон интернализуется внутрь клетки. К последней категории гормонов относится инсулин. Некоторые гормоны, такие как стероидные гормоны и гормоны щитовидной железы, по свойствам являются гидрофобными. В плазме крови для их транспортировки используются специальные белки-транспортеры. В комплексе с этими белками они не способны взаимодействовать с мембранными рецепторами, но способны отрываться от них и диффундировать через клеточную мембрану внутрь клетки. После перехода в цитозоль гормоны немедленно подхватываются другими белками, которые уже являются рецепторами. Комплекс гормон-рецептор в некоторых случаях дополнительно модифицируется и активируется. Далее он проникает в ядро, где может связываться с ядерным рецептором. В результате гормон приобретает сродство к ДНК. Связываясь с гормон-чувствительным элементом в ДНК, гормон влияет на транскрипцию определенных генов и изменяет концентрацию РНК в клетке и, соответственно, количество определенных белков в клетке.
27. Гормоны гипоталамуса – либерины и статины. Химическая природа. Механизм их действия на молекулярном уровне, биологический эффект.
Гипоталамус служит местом непосредственного взаимодействия высших отделов ЦНС и эндокринной системы. гипоталамуса были выделены первые гуморальные факторы, оказавшиеся гормональными веществами с чрезвычайно высокой биологической активностью. Эти вещества получили наименование рилизинг-факторов или либеринов. Вещества с противоположным действием, гипофизарных гормонов, стали называть ингибирующими факторами, или статинами. Таким образом, гормонам гипоталамуса принадлежит ключевая роль в физиологической системе гормональной регуляции многосторонних биологических функций отдельных органов, тканей и целостного организма. К настоящему времени в гипоталамусе открыто 7 стимуляторов (либерины) и 3 ингибитора (статины) секреции гормонов гипофиза, а именно: кортиколиберин, тиролиберин, люлиберин, фоллилиберин, соматолиберин, пролактолиберин, меланолиберин, соматостатин, пролактостатин и меланостатин. Установлено, что по химическому строению все гормоны гипоталамуса являются низкомолекулярными пептидами, так называемыми олигопептидами необычного строения, хотя точный аминокислотный состав и первичная структура выяснены не для всех. 1.Тиролиберин (Пиро-Глу–Гис–Про–NH2): представлен трипептидом, состоящим из пироглутаминовой (циклической) кислоты, гистидина и пролинамида, соединенных пептидными связями. В отличие от классических пептидов он не содержит свободных NH2- и СООН-групп у N- и С-концевых аминокислот. Ответственен за выпуск гормона, стимулирующего работу щитовидной железы. 2. Гонадолиберин является декапептидом, состоящим из 10 аминокислот в последовательности: Пиро-Глу–Гис–Трп–Сер–Тир–Гли–Лей–Арг–Про–Гли-NН2. Концевая С-аминокислота представлена глицинамидом. ГнРГ вызывает усиление секреции передней долей гипофиза гонадотропных гормонов —лютеинизирующего гормона и фолликулостимулирующего гормона. 3. Соматостатин является циклическим тетрадекапептидом (состоит из14 аминокислотных остатков). Отличается этот гормон от двух предыдущих, помимо циклической структуры, тем, что не содержит на N-конце пироглутаминовой кислоты: дисульфидная связь образуется между двумя остатками цистеина в 3-м и в 14-м положении. Он оказывает широкий спектрбиологического действия; в частности, показано ингибирующее действие насинтез гормона роста в аденогипофизе, а также прямое тормозящее действие его на биосинтез инсулина и глюкагона в β- и α-клетках островков Лангерганса. 4. Соматолиберин недавно выделен из природных источников. Он представлен 44 аминокислотными остатками с полностью раскрытой последовательностью. Биологической активностью соматолиберина наделен, кроме того, химически синтезированный декапептид: Н-Вал–Гис–Лей–Сер–Ала–Глу–Глн–Лиз–Глу–Ала-ОН. Этот декапептид стимулирует синтез и секрецию гормона роста гипофиза соматотропина. 5. Меланолиберин, химическая структура которого аналогична структуре открытого кольца гормона окситоцина (без трипептидной боковой цепи), имеет следующее строение: Н-Цис–Тир–Иле–Глн Асн–Цис-ОН. Необходимо отметить, что меланолиберин оказывает стимулирующее действие, а меланостатин, напротив, ингибирующее действие на синтез и секрецию меланотропина в передней доле гипофиза. 6. Меланостатин (меланотропинингибирующий фактор) представлен или трипептидом: Пиро-Глу–Лей–Гли-NН2, или пентапептидом со следующей последовательностью: Пиро-Глу–Гис–Фен–Aрг–Гли–NН2. МСГ стимулируют синтез и секрецию меланинов (меланогенез) клетками - меланоцитами кожи и волос, а также пигментного слоя сетчатки глаза. Наиболее сильное влияние на пигментацию оказывает альфа-меланоцитстимулирующий гормон.
28. Тропные гормоны гипофиза: соматотропный гормон, адренокортикотропный, тиреотропный, гонадотропные гормоны. Химическая природа. Механизм действия, биологический эффект. Нарушение функций организма и недостаточная или избыточная выработка соматотропина.
Адренокортикотропный гормон (АКТГ, кортикотропин) вырабатываемый базофильными клеткамиаденогипофиза. АКТГ, помимо основного действия – стимуляции синтезаи секреции гормонов коры надпочечников, обладает жиромобилизующейи меланоцитстимулирующей активностью. Молекула АКТГ содержит 39 аминокислотных остатков:
Н-Сер–Тир–Сер–Мет–Глу–Гис–Фен–Арг–Трп–Гли–Лиз–Про–Вал–Гли–
–Лиз–Лиз–Aрг–Aрг–Про–Вал–Лиз–Вал–Тир–Про–Асп–Ала–Гли–Глу–
–Асп–Глн–Сер–Ала–Глу–Ала–Фен–Про–Лей–Глу–Фен-ОН
Данные о механизме действия АКТГ на синтез стероидных гормонов свидетельствуют о существенной роли аденилатциклазной системы. Предполагают, что АКТГ вступает во взаимодействие со специфическими рецепторами на внешней поверхности клеточной мембраны (рецепторы представлены белками в комплексе с другими молекулами, в частности с сиаловой кислотой). Сигнал затем передается на фермент аденилатциклазу, расположенную на внутренней поверхности клеточной мембраны, которая катализирует распад АТФ и образование цАМФ. Последний активирует протеинкиназу, которая в свою очередь с участием АТФ осуществляет фосфорилирование холинэстеразы, превращающей эфиры холестерина в свободный холестерин, который поступает в митохондрии надпочечников, где содержатся все ферменты, катализирующие превращение холестерина в кортикостероиды. Соматотропный гормон (СТГ, гормон роста, соматотропин) синтезируется в ацидофильных клетках передней доли гипофиза; концентрация его в гипофизе составляет 5–15 мг на 1 г ткани. СТГ человека состоит из 191 аминокислоты и содержит две дисульфидные связи; N- и С-концевые аминокислоты представлены фенилаланином. СТГ обладает широким спектром биологического действия. Он влияет на все клетки организма, определяя интенсивность обмена углеводов, белков, липидов и минеральных веществ. Он усиливает биосинтез белка, ДНК, РНК и гликогена и в то же время способствует мобилизации жиров из депо и распаду высших жирных кислот и глюкозы в тканях. Помимо активации процессов ассимиляции, сопровождающихся увеличением размеров тела, ростом скелета, СТГ координирует и регулирует скорость протекания обменных процессов. Многие биологические эффекты этого гормона осуществляются через особый белковый фактор, образующийся в печени под влиянием гормона – соматомедин. По своей природе он оказался пептидом с мол. массой 8000. Тиреотропный гормон (ТТГ, тиротропин) является сложным гликопротеином и содержит, кроме того, по две α- и 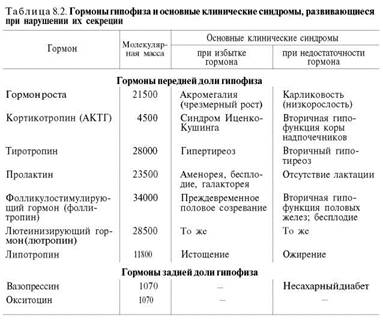 β-субъединицы,которые в отдельности биологической активностью не обладают: мол. масса его около 30000.Тиротропин контролирует развитие и функцию щитовидной железыи регулирует биосинтез и секрецию в кровь тиреоидных гормонов. Полностью расшифрована первичная структура α- и β-субъединиц тиротропина: α-субъединица, содержащая 96 аминокислотныхостатков; β-субъединица тиротропина человека, содержащая 112 аминокислотных остатков, К гонадотропным гормонам (гонадотропины) относятся фолликулостимулирующий гормон (ФСГ,фоллитропин) и лютеинизирующий гормон (ЛГ, лютропин). Оба гормона синтезируютсяв передней доле гипофиза и являются сложнымибелками – гликопротеинами с мол. массой 25000. Они регулируют стероидо - и гаметогенез в половых железах. Фоллитропин вызывает созревание фолликулов в яичниках у самок и сперматогенез – у самцов. Лютропину самок стимулирует секрецию эстрогенов и прогестерона, как и разрыв фолликулов с образованием желтого тела, а у самцов – секрецию тестостерона и развитие интерстициальной ткани. Биосинтез гонадотропинов,как было отмечено, регулируется гипоталамическим гормоном гонадолиберином.Лютропин состоит из двух α- и β-субъединиц: α-субъединица гормона содержит из 89 аминокислотных остатков с N-конца и отличается природой 22 аминокислот.
β-субъединицы,которые в отдельности биологической активностью не обладают: мол. масса его около 30000.Тиротропин контролирует развитие и функцию щитовидной железыи регулирует биосинтез и секрецию в кровь тиреоидных гормонов. Полностью расшифрована первичная структура α- и β-субъединиц тиротропина: α-субъединица, содержащая 96 аминокислотныхостатков; β-субъединица тиротропина человека, содержащая 112 аминокислотных остатков, К гонадотропным гормонам (гонадотропины) относятся фолликулостимулирующий гормон (ФСГ,фоллитропин) и лютеинизирующий гормон (ЛГ, лютропин). Оба гормона синтезируютсяв передней доле гипофиза и являются сложнымибелками – гликопротеинами с мол. массой 25000. Они регулируют стероидо - и гаметогенез в половых железах. Фоллитропин вызывает созревание фолликулов в яичниках у самок и сперматогенез – у самцов. Лютропину самок стимулирует секрецию эстрогенов и прогестерона, как и разрыв фолликулов с образованием желтого тела, а у самцов – секрецию тестостерона и развитие интерстициальной ткани. Биосинтез гонадотропинов,как было отмечено, регулируется гипоталамическим гормоном гонадолиберином.Лютропин состоит из двух α- и β-субъединиц: α-субъединица гормона содержит из 89 аминокислотных остатков с N-конца и отличается природой 22 аминокислот.
29. Гормоны задней доли гипофиза: вазопрессин, окситоцин. Химическая природа. Механизм их действия, биологический эффект. Нарушения функций организма, связанные с недостатком выработки этих гормонов.
Гормоны вазопрессин и окситоцин синтезируются рибосомальным путем. Оба гормона представляют собой нонапептиды следующего строения: Вазопрессин отличается от окситоцина двумя аминокислотами: он содержит в положении 3 от N-конца фенилаланин вместо изолейцина и в положении 8 – аргинин вместо лейцина. Основной биологический эффект окситоцина у млекопитающих связан со стимуляцией сокращения гладких мышц матки при родах и мышечных волокон вокруг альвеол молочных желез, что вызывает секрецию молока. Вазопрессин стимулирует сокращение гладких мышечных волокон сосудов, оказывая сильное вазопрессорное действие, однако основная роль его в организме сводится к регуляции водного обмена, откуда его второе название антидиуретического гормона. В небольших концентрациях (0,2 нг на 1 кг массы тела) вазопрессин оказывает мощное антидиуретическое действие – стимулирует обратный ток воды через мембраны почечных канальцев. В норме он контролирует осмотическое давление плазмы крови и водный баланс организма человека. При патологии, в частности атрофии задней доли гипофиза, развивается несахарный диабет – заболевание, характеризующееся выделением чрезвычайно больших количеств жидкости с мочой. При этом нарушен обратный процесс всасывания воды в канальцах почек.
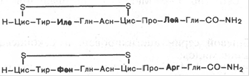 Окситоцин
Окситоцин
Вазопрессин
30. Гормоны щитовидной железы: трийодтиронин и тироксин. Химическая природа, биосинтез. Механизм действия гормонов на молекулярном уровне, биологический эффект. Изменение обмена вещества при гипертиреозе. Механизм возникновения эндемического зоба и его предупреждение.
Тироксин и трийодтиронин – основные гормоны фолликулярной части щитовидной железы. Помимо этих гормонов (биосинтез и функции которых будут рассмотрены ниже), в особых клетках – так называемых парафолликулярных клетках, или С-клетках щитовидной железы, синтезируется гормон пептидной природы, обеспечивающий постоянную концентрацию кальция в крови. Он получил название ≪ кальцитонин ≫. Биологическое действие кальцитонина прямо противоположно эффекту паратгормона: он вызывает подавление в костной ткани резорбтивных процессов и соответственно гипокальциемию и гипофосфатемию. Из L-тиронина легко синтезируется гормон щитовидной железы тироксин, содержащий в 4 положениях кольцевой структуры йод Биологическое действие гормонов щитовидной железы распространяется на множество физиологических функций организма. В частности, гормоны регулируют скорость основного обмена, рост и дифференцировку тканей, обмен белков, углеводов и липидов, водно-электролитный обмен, деятельность ЦНС, пищеварительного тракта, гемопоэз, функцию сердечнососудистой системы, потребность в витаминах, сопротивляемость организма инфекциям и др. Гипофункция щитовидной железы в раннем детском возрасте приводит к развитию болезни, известной в литературе как кретинизм. Помимо остановки роста, специфических изменений кожи, волос, мышц, резкого снижения скорости процессов обмена, при кретинизме отмечаются глубокие нарушения психики; специфическое гормональное лечение в этом случае не дает положительных результатов. Повышенная функция щитовидной железы (гиперфункция) вызывает развитие гипертиреоза
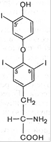
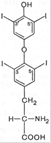
L-тироксин L-3,5,3'-трийодтиронин
31. Гормоны коры надпочечников: глюкокортикоиды, минералокортикоиды. Химическая природа. Механизм действия на молекулярном уровне. Их роль в регуляции углеводного, минерального, липидного и белкового обмена.
В зависимости от характера биологического эффекта гормоны коркового вещества надпочечников условно делят на глюкокортикоиды (кортикостероиды, оказывающие влияние на обмен углеводов, белков, жиров и нуклеиновых кислот) и минералокортикоиды (кортикостероиды, оказывающие преимущественное влияние на обмен солей и воды). К первым относятся кортикостерон, кортизон, гидрокортизон (кортизол), 11-дезоксикортизол и 11-дегидрокортикостерон, ко вторым – дезоксикортикостерон и альдостерон. В основе их структуры, так же как и в основе строения холестерина, эргостерина, желчных кислот, витаминов группы D, половых гормонов и ряда других веществ, лежит конденсированная кольцевая система циклопентанпергидрофенантрена. Глюкокортикоиды оказывают разностороннее влияние на обмен веществ в разных тканях. В мышечной, лимфатической, соединительной и жировой тканях глюкокортикоиды, проявляя катаболическое действие, вызывают снижение проницаемости клеточных мембран и соответственно торможение поглощения глюкозы и аминокислот; в то же время в печени они оказывают противоположное действие. Конечным итогом воздействия глюкокортикоидов является развитие гипергликемии, обусловленной главным образом глюконеогенезом. Минералокортикоиды (дезоксикортикостерон и альдостерон) регулируют главным образом обмен натрия, калия, хлора и воды; они способствуют удержанию ионов натрия и хлора в организме и выведению с мочой ионов калия. По-видимому, происходит обратное всасывание ионов натрия и хлора в канальцах почек в обмен на выведение других продуктов обмена,
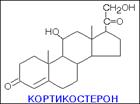
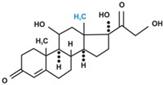
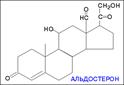
кортизол
32. Паратгормон и кальцитонин. Химическая природа. Механизм действия на молекулярном уровне. Влияние на обмен кальция, гиперкальциемия и гипокальциемия.
К гормонам белковой природы относится также паратиреоидный гормон (паратгормон). Они синтезируются паращитовидными железами. Молекула паратгормона быка содержит 84 аминокислотных остатка и состоит из одной полипептидной цепи. Выяснено, что паратгормон участвует в регуляции концентрации катионов кальция и связанных с ними анионов фосфорной кислоты в крови. Биологически активной формой считается ионизированный кальций, концентрация его колеблется в пределах 1,1–1,3 ммоль/л. Ионы кальция оказались эссенциальными факторами, не заменимыми другими катионами для ряда жизненно важных физиологических процессов: мышечное сокращение, нервно-мышечное возбуждение, свертывание крови, проницаемость клеточных мембран, активность ряда ферментов и т.д. Поэтому любые изменения этих процессов, обусловленные длительным недостатком кальция в пище или нарушением его всасывания в кишечнике, приводят к усилению синтеза паратгормона, который способствует вымыванию солей кальция (в виде цитратов и фосфатов) из костной ткани и соответственно к деструкции минеральных и органических компонентов костей. Другой орган-мишень паратгормона – это почка. Паратгормон уменьшает реабсорбцию фосфата в дистальных канальцах почки и повышает канальцевую реабсорбцию кальция. В особых клетках – так называемых парафолликулярных клетках, или С-клетках щитовидной железы, синтезируется гормон пептидной природы, обеспечивающий постоянную концентрацию кальция в крови - кальцитонин. Формула:
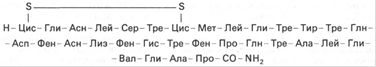
Кальцитонин содержит дисульфидный мостик (между 1-м и 7-м аминокислотными остатками) и характеризуется N-концевым цистеином и С-концевым пролинамидом. Биологическое действие кальцитонина прямо противоположно эффекту паратгормона: он вызывает подавление в костной ткани резорбтивных процессов и соответственно гипокальциемию и гипофосфатемию. Таким образом, постоянство уровня кальция в крови человека и животных обеспечивается главным образом паратгормоном, кальцитриолом и кальцитонином, т.е. гормонами как щитовидной и паращитовидных желез, так и гормоном – производным витамина D3. Это следует учитывать при хирургических лечебных манипуляциях на данных железах.
33. Гормоны мозгового слоя надпочечников – катехоламины: адреналин и норадреналин. Химическая природа и биосинтез. Механизм действия гормонов на молекулярном уровне, их роль в регуляции обмена углеводов, жиров и аминокислот. Нарушения обмена при заболеваниях надпочечников.
Эти гормоны по строению напоминают аминокислоту тирозин, от которого они отличаются наличием дополнительных ОН-групп в кольце и у β-углеродного атома боковой цепи и отсутствием карбоксильной группы.
Адреналин Норадреналин Изопропиладреналин
В мозговом веществе надпочечников человека массой 10 г содержится около 5 мг адреналина и 0,5 мг норадреналина. Содержание их в крови составляет соответственно 1,9 и 5,2 нмоль/л. В плазме крови оба гормона присутствуют как в свободном, так и в связанном, в частности, с альбуминами состоянии. Небольшие количества обоих гормонов откладываются в виде соли с АТФ в нервных окончаниях, освобождаясь в ответ на ихраздражение. Кроме того, все они о казывают мощное сосудосуживающее действие, вызывая повышение артериального давления, и в этом отношении действие их сходно с действием симпатической нервной системы. Известно мощное регулирующее влияние этихгормонов на обмен углеводов в организме. Так, в частности, адреналинвызывает резкое повышение уровня глюкозы в крови, что обусловленоускорением распада гликогена в печени под действием фермента фосфорилазы. Гипергликемическйй эффект норадреналина значительно ниже – примерно 5% от действия адреналина. Параллельно отмечаются накопление гексозофосфатов в тканях, в частности в мышцах, уменьшение концентрации неорганического фосфата и повышение уровня ненасыщенных жирных кислот в плазме крови. Имеются данные о торможении окисления глюкозы в тканях под влиянием адреналина. Это действие некоторые авторы связывают с уменьшением скорости проникновения (транспорта) глюкозы внутрь клетки. Известно, что и адреналин, и норадреналин быстро разрушаются в организме; с мочой выделяются неактивные продукты их обмена, главным образом в виде 3-метокси-4-оксиминдальной кислоты, оксоадренохрома, метоксинорадреналина и метоксиадреналина. Эти метаболиты содержатся в моче преимущественно в связанной с глюкуроновой кислотой форме. Ферменты, катализирующие указанные превращения катехоламинов, выделены из многих тканей и достаточно хорошо изучены, в частности моноаминоксидаза (МАО), определяющая скорость биосинтеза и распада катехоламинов, и катехолметилтрансфераза, катализирующая главный путь превращения адреналина, т.е. о- метилирование за счет S-аденозилметионина. Приводим структуру двух конечных продуктов распада
34. Глюкагон и инсулин. Химическая природа, биосинтез инсулина. Механизм действия этих гормонов на молекулярном уровне. Их роль в регуляции обмена углеводов, жиров, аминокислот. Биохимические нарушения при сахарном диабете.
 Инсулин, получивший свое название от наименования панкреатических островков. Молекула инсулина, содержащая 51 аминокислотный остаток, состоит из двух полипептидных цепей, соединенных между собой в двух точках дисульфидными мостиками. В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в крови. Так, повышение содержания глюкозы в крови вызывает увеличение секреции инсулина в панкреатических островках, а снижение ее содержания, наоборот,– замедление секреции инсулина. Этот феномен контроля по типу обратной связи рассматривается как один из важнейших механизмов регуляции содержания глюкозы в крови. При недостаточной секреции инсулина развивается специфическое заболевание – сахарный диабет. Физиологические эффекты инсулина: Инсулин — единственный гормон, снижающий содержание глюкозы в крови, это реализуется через:
Инсулин, получивший свое название от наименования панкреатических островков. Молекула инсулина, содержащая 51 аминокислотный остаток, состоит из двух полипептидных цепей, соединенных между собой в двух точках дисульфидными мостиками. В физиологической регуляции синтеза инсулина доминирующую роль играет концентрация глюкозы в крови. Так, повышение содержания глюкозы в крови вызывает увеличение секреции инсулина в панкреатических островках, а снижение ее содержания, наоборот,– замедление секреции инсулина. Этот феномен контроля по типу обратной связи рассматривается как один из важнейших механизмов регуляции содержания глюкозы в крови. При недостаточной секреции инсулина развивается специфическое заболевание – сахарный диабет. Физиологические эффекты инсулина: Инсулин — единственный гормон, снижающий содержание глюкозы в крови, это реализуется через:
§ усиление поглощения клетками глюкозы и других веществ;
§ активацию ключевых ферментов гликолиза;
§ увеличение интенсивности синтеза гликогена — инсулин форсирует запасание глюкозы клетками печени и мышц путём полимеризации её в гликоген;
§ уменьшение интенсивности глюконеогенеза — снижается образование в печени глюкозы из различных веществ
Анаболические эффекты
§ усиливает поглощение клетками аминокислот (особенно лейцина и валина);
§ усиливает транспорт в клетку ионов калия, а также магния и фосфата;
§ усиливает репликацию ДНК и биосинтез белка;
§ усиливает синтез жирных кислот и последующую их этерификацию — в жировой ткани и в печени инсулин способствует превращению глюкозы в триглицериды; при недостатке инсулина происходит обратное — мобилизация жиров.
Антикатаболические эффекты
§ подавляет гидролиз белков — уменьшает деградацию белков;
§ уменьшает липолиз — снижает поступление жирных кислот в кровь.
Глюкагон — гормон альфа-клеток островков Лангерганса поджелудочной железы. По химическому строению глюкагон является пептидным гормоном. Молекула глюкагона состоит из 29 аминокислот и имеет молекулярный вес 3485. Первичная структура молекулы глюкагона следующая:
NH2-His-Ser-Gln-Gly-Thr-Phe- Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser- Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp-Leu- Met-Asn-Thr-COOH
Механизм действия глюкагона обусловлен его связыванием со специфическими глюкагоновыми рецепторами клеток печени. Это приводит к повышению опосредованной G-белком активности аденилатциклазы и увеличению образования цАМФ. Результатом является усиление катаболизма депонированного в печени гликогена. Глюкагон оказывает сильное инотропное и хронотропное действие на миокард вследствие увеличения образования. Результатом является повышение артериального давления, увеличение частоты и силы сердечных сокращений
35. Половые гормоны: андрогены, эстрогены. Химическая природа. Механизм действия на молекулярном уровне, влияние на обмен веществ. Нарушения обмена при недостатке и избытке этих гормонов в организме.
Женские половые гормоны. Основным местом синтеза женских половых гормонов – эстрогенов – являются яичники и желтое тело. В настоящее время открыты 2 группы женских половых гормонов, различающихся своей химической структурой и биологической функцией: эстрогены (главный представитель – эстрадиол) и прогестины (главный представитель – прогестерон). Наиболее активный эстроген – эстрадиол, синтезируется преимущественно в фолликулах; два остальных эстрогена являются производными эстрадиола и синтезируются также в надпочечниках и плаценте. Все эстрогены состоят из 18 атомов углерода. Секреция эстрогенов и прогестерона яичником носит циклический характер, зависящий от фазы полового цикла: в первой фазе цикла синтезируются в основном эстрогены, а во второй – преимущественно прогестерон. Основная биологическая роль эстрогенов и прогестерона, синтез которых начинается после наступления половой зрелости, заключается в обеспечении репродуктивной функции организма женщины. В этот период они вызывают развитие вторичных половых признаков и создают оптимальные условия, обеспечивающие возможность оплодотворения яйцеклетки после овуляции. Прогестерон выполняет в организме ряд специфических функций: подготавливает слизистую оболочку матки к успешной имплантации яйцеклетки в случае ее оплодотворения, а при наступлении беременности основная роль – сохранение беременности; оказывает тормозящее влияние на овуляцию и стимулирует развитие ткани молочной железы. Эстрогены оказывают анаболическое действие на организм, стимулируя синтез белка. Мужские половые гормоны. Биосинтез андрогенов осуществляется главным образом в семенникахи частично в яичниках и надпочечниках Биологическая роль андрогенов в мужском организме в основном связана с дифференцировкой и функционированием репродуктивной системы, причем в отличие от эстрогенов андрогенные гормоны уже в эмбриональном периоде оказывают существенное влияние на дифференцировку мужских половых желез, а также других тканей, определяя характер секреции гонадотропных гормонов у взрослых. Во взрослом организме андрогены регулируют развитие мужских вторичных половых признаков, сперматогенез в семенниках и т.д.
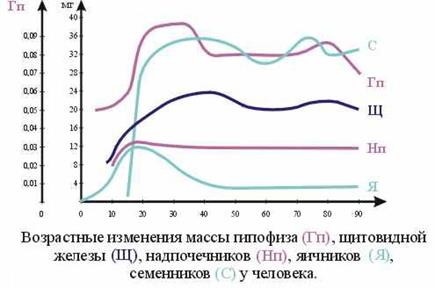
36. Неравномерность созревания и функционирования желез внутренней секреции у ребенка в различные возрастные периоды. Отличие общего гормонального фона ребенка по сравнению с организмом взрослых.
37. Мембраны, их значение для клеток и клеточных включений. Липидный бислой как основа плазматических и других клеточных мембран. Однотипичность их строения. Гликолипиды и белки мембран. Физико-химические свойства мембраны. Липидный и фосфолипидный спектр мембраны. ПОЛ – важный физиологический процесс. Изменения ПОЛ при патологии, пути защиты мембраны от ПОЛ.
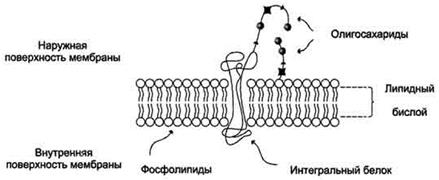
Клеточная мембрана отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулируют обмен между клеткой и средой; внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки — компартменты или органеллы, в которых поддерживаются определённые условия среды. Биологические мембраны представляют собой "ансамбли" липидных и белковых молекул, удерживаемых вместе с помощью нековалентных взаимодействий. Основу мембраны составляет двойной липидный слой, в формировании которого участвуют фосфолипиды и гликолипиды. Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы "растворены" в липидном бислое. Фосфолипиды. Все фосфолипиды можно разделить на 2 группы - глицерофосфолипиды и сфингофосфолипиды. Глицерофосфолипиды относят к производным фосфатидной кислоты. Наиболее распространённые глицерофосфолипиды мембран - фосфатидилхолины и фосфатидилэтаноламины. В мембранах эукариотических клеток обнаружено огромное количество разных фосфолипидов, причём они распределены неравномерно по разным клеточным мембранам. Эта неравномерность относится к распределению как полярных "головок", так и ацильных остатков. Г ликолипиды. В гликолипидах гидрофобная часть представлена церамидом. Гидрофильная группа - углеводный остаток, присоединённый гликозидной связью к гидроксильной группе у первого углеродного атома церамида. В зависимости от длины и строения углеводной части различают цереброзиды, содержащие моно - или олигосахаридный остаток, и ганглиозиды, к ОН-группе которых присоединён сложный, разветвлённый олигосахарид, содержащий N-ацетилнейраминовую кислоту (NANA). Специфические свойства биологических мембран. Благодаря указанным особенностям биологические мембраны имеют присущие им характерные черты. Они образуют протяженные бислойные структуры малой толщины (6–10 нм), объединяющие белковые и липидные компоненты с различными свойствами. Целостная структура мембраны создается за счет гидрофобных и электростатических взаимодействий, а не за счет ковалентных связей между составляющими ее молекулами белков и липидов. Гидрофобный липидный бислой представляет естественную преграду для проникновения полярных молекул. Мембраны асимметричны по своему исходному строению, что обеспечивает градиент кривизны и спонтанное образование замкнутых структур. Мембранный бислой обладает относительно малой микровязкостью. Другими словами, мембраны рыхло упакованы, что позволяет отдельным компонентам проявлять высокую подвижность в латеральном направлении. Наружные мембраны клеток отличаются от внутренних по липидному составу (последние почти не содержат стеринов, имеют соотношение ФХ/ФЭ > 1) и обладают специфическим набором ферментов и рецепторов.
Как правило, белки плазматических мембран со стороны внеклеточной среды обильно гликозилированы. Внутриклеточные мембраны содержат мало гликопротеинов и гликолипидов и характеризуются меньшей микро- вязкостью. Благодаря этому они могут образовывать органеллы малого размера. Мембранные белки выполняют различные специфические функции: рецепторные, транспортные, ферментативные, энергопреобразующие и т.д.
38. Функции мембраны: барьерная, защитная, регуляторная, энергетическая. Транспортная функция мембраны. Виды транспорта: пассивный, активный, цитоз. Ферментный спектр мембраны: транспортные АТФ-азы, механизм их действия. Повреждение мембраны – неизбежный этап любой клеточной патологии. Атеросклероз
· барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
· матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие.
· механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
· энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
· рецепторная — некоторые белки, находящиеся в мембране, являются рецепторами.
ферментативная — мембранные белки нередко являются ферментами.
· Транспортная функцияявляется одной из важных функций клеточных мембран. Мембрана создает
существенные ограничения для проникновения различных веществ, однако она не является полностью непроницаемой: небольшие нейтральные молекулы могут проникать через бислой в области структурных дефектов. Этот процесс осуществляется по градиенту концентрации переносимого вещества - из области, где его содержание высоко, в область с более низким содержанием. Такой процесс называется простой диффузией, он осуществляется неизбирательно и с низкой скоростью. При облегченной диффузии вещества также переносятся в направлении их концентрационного градиента, но с использованием специальных структур - переносчиков или каналов, увеличивающих скорость и специфичность переноса. Известны высокоспецифические транслоказы - белковые молекулы, переносящие адениловые нуклеотиды через внутреннюю мембрану митохондрий: Nа+/Са2+-обменник - белок, входящий в состав плазматических мембран многих клеток; низкомолекулярный пептид бактериального происхождения валиномицин - специфический переносчик для ионов К+. Активный транспорт веществ осуществляется такими же механизмами, но протекает против концентрационного градиента и для своего осуществления должен быть сопряжен с энергодающим процессом. Основным источником энергии для активно- го транспорта является АТФ. Поэтому, как правило, эти системы представляют собой АТФазы. Примером систем активного транспорта ионов является Na+/K+- АТФаза плазматических мембран животных клеток, которая ≪выкачивает≫ из клетки ионы натрия в обмен на ионы калия, затрачивая на выполнение этой работы АТФ в стехиометрии 3Nа+/2К+/1АТФ. Са2+-АТФаза осуществляет активный транспорт кальция через мембрану со стехиометрией 2Са2+/1АТФ. В так называемых сопрягающих мембранах имеются протонные насосы, работающие как Н+-АТФазы. В результате их функционирования на мембране возникает разность концентраций протонов (ΔрН) и разность электрических потенциалов. Атеросклероз — хроническое заболевание артерий эластического и мышечно-эластического типа, возникающее вследствие нарушения липидного обмена и сопровождающееся отложением холестерина и некоторых фракций липопротеидов в интиме сосудов. Отложения формируются в виде атероматозных бляшек. Последующее разрастание в них соединительной ткани (склероз), и кальциноз стенки сосуда приводят к деформации и сужению просвета вплоть до облитерации (закупорки). Атеросклероз сосудов ведет к развитию ишемической болезни сердца.
39. Слабость антиоксидантной защиты у мембран у детей ранних сроков жизни и ее последующая активация.
40. Окислительное декарбоксилирование пировиноградной кислоты: последовательность реакций, строение пируватдегидрогеназного комплекса. Нарушение его функции при атеросклерозе
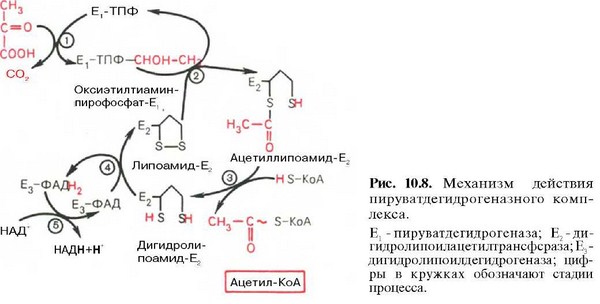
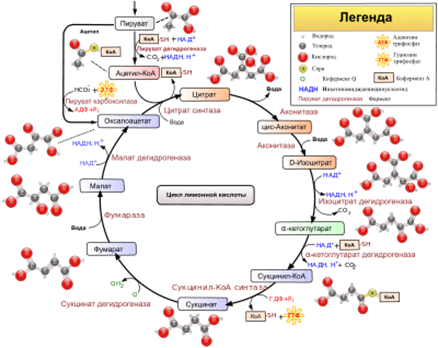
41. Цикл лимонной кислоты: последовательность реакций и характеристика ферментов. Связь между циклом трикарбоновых кислот и цепью переноса электронов и протонов. Аллостерическая регуляция цитратного цикла, его анаболические функции.
42. Понятие об обмене веществ и метаболических путях. Стадии катаболизма, взаимосвязь их с анаболическими процессами. Основные конечные продукты метаболизма у человека.
Метаболизм или обмен веществ — набор химических реакций, которые возникают в живом организме для поддержания жизни. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды. Метаболизм обычно делят на две стадии: в ходе катаболизма сложные органические вещества деградируют до более простых; в процессах анаболизма с затратами энергии синтезируются такие вещества, как белки, сахара, липиды и нуклеиновые кислоты. Обмен веществ происходит между клетками организма и межклеточной жидкостью, постоянство состава которой поддерживается кровообращением: за время прохождения крови в капиллярах через проницаемые стенки капилляров плазма крови 40 раз полностью обновляется с интерстициальной жидкостью. Серии химических реакций обмена веществ называют метаболическими путями, в них при участии ферментов одни биологически значимые молекулы последовательно превращаются в другие. Основные метаболические пути и их компоненты одинаковы для многих видов. Например, некоторые карбоновые кислоты, являющиеся интермедиатами цикла трикарбоновых кислот присутствуют во всех организмах Катаболические, анаболические пути. Ферментативное расщепление основных питательных веществ, а именно углеводов, жиров и белков, происходит в клетке через ряд последовательных ферментативных реакций. Катаболизм основных питательных веществ включает три главные стадии. На первой стадии крупные пищевые молекулы расщепляются на составляющие их основные строительные блоки. Полисахариды, например, расщепляются до гексоз или пентоз, липиды — до жирных кислот, глицерина и других компонентов, белки — до аминокислот, которых имеется 20 видов. На второй стадии большое число продуктов, образовавшихся на первой стадии, превращается в более простые молекулы, число типов которых сравнительно невелико. Так, гексозы, пентозы и глицерин, разрушаясь, превращаются сначала в трехуглеродный фосфорилированный сахар — глицеральдегид-3-фосфат, а затем расщепляются далее до единственной двууглеродной формы — ацетильной группы, входящей в состав ацетил- КоА. Двадцать различных аминокислот также дают при расщеплении лишь несколько конечных продуктов, а именно ацетил-КоА, α-кетоглутарат, сукцинат, фумарат и оксалоацетат. Продукты, образовавшиеся на второй стадии, вступают в третью стадию, которая для них всех является общей и на которой они в конечном счете окисляются до CO2 и воды. Процесс анаболизма также включает три стадии. Исходными веществами, или строительными блоками служат для него соединения, поставляемые третьей стадией катаболизма. Таким образом, третья стадия катаболизма является в то же время первой, исходной, стадией анаболизма. Синтез белка, например, начинается на этой стадии с α - кетокислот, являющихся предшественниками α -аминокислот. На второй стадии анаболизма α -кетокислоты аминируются аминогруппой доноров с образованием α -аминокислот, а на третьей, заключительной, стадии аминокислоты объединяются в пептидные цепи. Продукты метаболизма выводятся с мочой, фекалиями, выдыхаемым воздухом и потом. Специфические вещества задерживаются или удаляются из организма в той степени, в какой это необходимо для поддержания гомеостаза, при этом вместе со шлаковыми продуктами распада выводятся потенциально полезные вещества. Небольшие количества этих веществ выводятся из организма в виде кишечных газов, волос, ногтей, десквамированного эпителия кожи, кожного сала, ушной серы, слизи из полости носа и влагалища, слюны, слез, семенной жидкости и менструальных выделений. Уровни потерь этих веществ опубликованы в работах.
43. Митохондриальное окисление. Дегидрирование субстратов и окисление водорода как источник энергии для синтеза АТФ. Структура и функции переноса электронов, окислительно-восстановительный потенциал структурных компонентов цепи. Нарушение функции цепи переноса электронов при гипоксиях и других патологических состояниях.
Система митохондриального окисления - мультиферментная система, постепенно транспортирующая протоны и электроны на кислород с образованием молекулы воды. Все ферменты митохондриального окисления встроены во внутреннюю мембрану митохондрий. Только первый переносчик протонов и электронов - никотинамидная дегидрогеназа расположена в матриксе митохондрии. Этот фермент отнимает водород от субстрата и передает его следующему переносчику. Полный комплекс таких ферментов образует «дыхательную цепь», в пределах которого атомы водорода отнимаются от субстрата, затем передаются последовательно от одного переносчика к другому, и, наконец, передаются на кислород воздуха с образованием воды. Существует строгая последовательность работы каждого звена в цепочке переносчиков. Эта последовательность определяется величиной редокс-потенциала (окислительно-восстановительного потенциала, сокращенно - овп) каждого звена. ОВП - это химическая характеристика способности вещества принимать и удерживать электроны. Вещества с положительным ОВП окисляют водород (отнимают от него электроны), вещества с отрицательным ОВП окисляются самим водородом. Самый низкий ОВП имеет начальное звено цепи, самый высокий - у кислорода, расположенного в конце цепочки переносчиков. Таким образом, передача водорода идет от более низкого к более высокому ОВП. На одной из стадий происходит разделение атомов водорода на Н+ и электроны. Протоны остаются временно в окружающей среде, а электроны идут дальше по цепи и в ее конце используются для активации О2. Кислород является конечным акцептором электронов. O2 + 4e -----> 2O-2 (полное восстановление кислорода). Все реакции, происходящие в дыхательной цепи, сопряжены. Переносчики водорода и электронов расположены в строгом порядке, в соответствии с величиной их редокс-потенциала. В настоящее время различают три варианта дыхательных цепей: Главная дыхательная цепь - это три мультиферментных комплекса, встроенных во внутреннюю мембрану митохондрии. Обозначаются они латинскими цифрами – I, III и IV. Сокращенная (укороченная) дыхательная цепи. Известны два субстрата. Главным из них является сукцинат (янтарная кислота). Сукцинатдегидрогеназа – это комплекс II, который в укороченном варианте цепи является начальным звеном окисления. В составе комплекса – простетическая группа ФАД и FeSII. От ФАД.Н2 два атома водорода переносятся на KoQ. Значит, первое звено, которое имеется в полной цепи - исключается. Перепад окислительно-восстановительного потенциала между ФАД и KoQ невелик. Поэтому переноса H+ в межмембранное пространство в этой точке не происходит. DmH+ также создается, но меньший, чем в полной цепи. Значит, меньше и эффективность фосфорилирования - коэффициент Р/О=2. Аналогичным образом окисляется и второй субстрат – ацил-КоА (активная форма любой жирной кислоты): Максимально сокращенная (максимально укороченная) дыхательная цепь. Она представлена только цитохромной частью. Эксперименты показали, что здесь может быть окислен только один субстрат - аскорбиновая кислота, с участием фермента, восстановленные эквиваленты включаются в цепь на уровне цитохрома С цитохромоксидазы (цитохром аа3), но в реальных условиях такого окисления практически не происходит. Образуется вода и 1 молекула АТФ. Коэффициент Р/О=1.
44. Окислительное фосфорилирование, его сопряжение с дыхательной цепью. Химиоосмотическая теория Митчела. Коэффициент Р/О. Разобщение тканевого дыхания и окислительного фосфорилирования. Биологические разобщители как регуляторы окислительного фосфорилирования. Нарушения энергетического обмена: гипоксические состояния.
Окислительное фосфорилирование и дыхательный контроль. Функция дыхательной цепи – утилизация восстановленных дыхательных переносчиков, образующихся в реакциях мета








